
Митохондриальный каскад при спорадической форме болезни Альцгеймера в эксперименте
Поздняков Д.И. Митохондриальный каскад при спорадической форме болезни Альцгеймера в эксперименте. Лабораторные животные для научных исследований. 2022; 2. https://doi.org/10.29296/2618723X-2022-02-06
Резюме
Патофизиологические реакции митохондриального каскада являются значимой составляющей патогенеза нейродегенеративных заболеваний, в частности болезни Альцгеймера, и могут служить базисом для нейропротекторного вмешательства, что предполагает актуальность их изучения.
Цель исследования. Оценить зависимость изменения показателей митохондриальной функции от меняющегося содержания тау-белка в гиппокампе крыс в условиях экспериментальной болезни Альцгеймера спорадического типа.
Материал и методы. Болезнь Альцгеймера моделировали у крыс Wistar путем введения фрагментов Аβ1–42 в СА1‑часть гиппокампа. Через 60 дней экспозиции у крыс оценивали развитие когнитивного дефицита в тесте Y-образного лабиринта. Когнитивный дефицит у животных оценивали по изменению числа спонтанных чередований рукавов лабиринта. В гиппокампе животных оценивали концентрацию АТФ, тау-протеина, митохондриального пероксида водорода и активность ферментов: аконитазы, цитратсинтазы, цитохром‑с-оксидазы, сукцинатдегидрогеназы. Также регистрировали величину митохондриального мембранного потенциала. Содержание АТФ и тау-белка определяли методом иммуноферментного анализа. Активность митохондриальных ферментов оценивали спектрометрически при внесении в реакционную среду соответствующих субстратов. Концентрацию митохондриального пероксида водорода определяли по изменению флюоресценции резоруфина. Мембранный потенциал митохондрий оценивали спектрофотометрическим методом. Данные обрабатывали статистически. В ходе анализа рассчитывали коэффициент корреляции Спирмена.
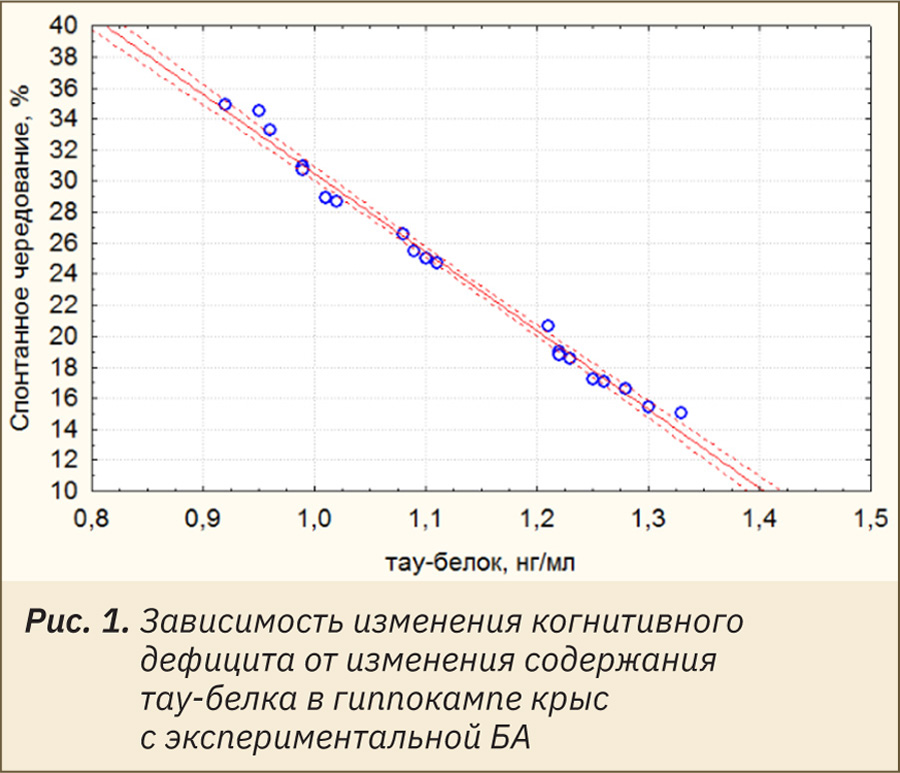
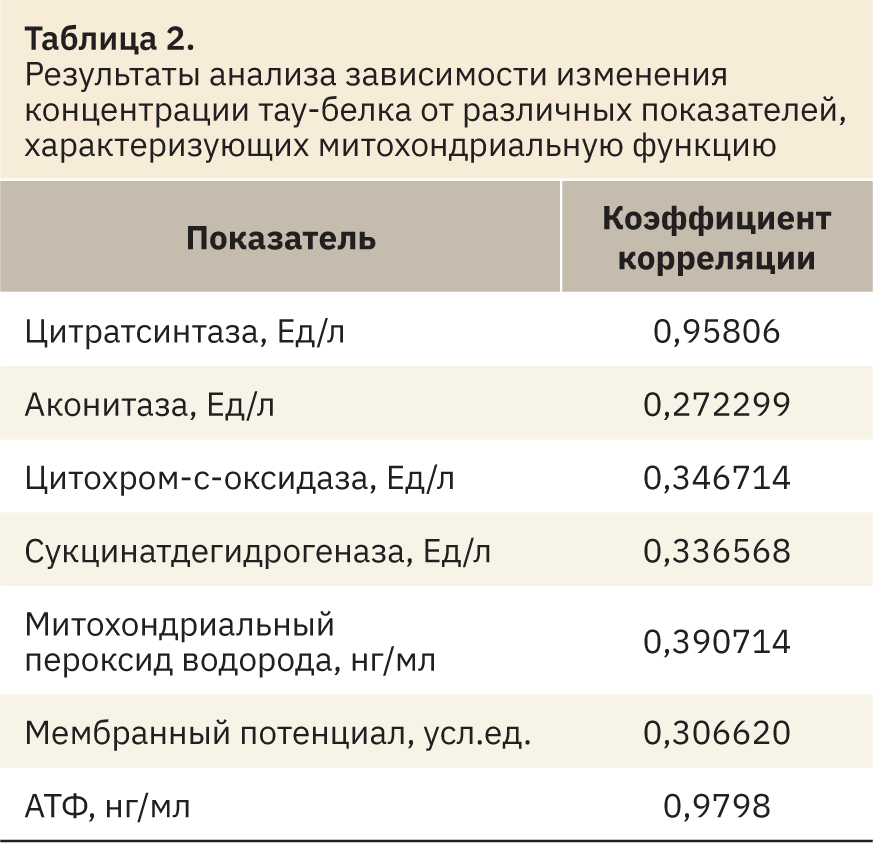
Результаты. В ходе исследования установлено, что у крыс с болезнью Альцгеймера по сравнению с ложнооперированными животными отмечено достоверное снижение активности цитратсинтазы, аконитазы, цитохром‑с-оксидазы, сукцинатдегидрогеназы, митохондриального мембранного потенциала и концентрации АТФ при повышении содержания митохондриального пероксида водорода и тау-белка. Также у крыс с экспериментальной патологией наблюдалось развитие выраженного когнитивного дефицита, прямо коррелирующего с концентрацией тау-белка (r=0,9950). В свою очередь изменение содержания тау-протеина коррелировало с различной активностью цитратсинтазы (r=0,95806) и концентрацией АТФ (r=0,9798). Полученные данные могут свидетельствовать, что накопление агрегатов фосфорилированного тау-белка в ткани головного мозга при болезни Альцгеймера может зависеть от митохондриального биогенеза (активность цитратсинтазы) и интегральной активности митохондрий клетки (концентрация АТФ).
Введение
Болезнь Альцгеймера (БА) — одна из самых распространенных терминальных форм деменции, ежегодно регистрирующаяся более чем у 4,5 млн человек. На сегодняшний день, согласно данным Всемирной организации здравоохранения (ВОЗ), БА стоит на 7‑м месте в списке ведущих причин смертности, а в экономически развитых странах данное заболевание по числу летальных исходов уступает лишь ишемической болезни сердца, занимая 2‑е место [1]. Во многом высокое медицинское, социальное и экономическое бремя БА побуждает научное сообщество не только к изучению особенностей этиопатогенеза заболевания, но и к разработке новых методов лечения. За последнее 10‑летие был достигнут существенный прогресс в понимании патогенеза БА. Основной гипотезой, которая объясняет нейродегенеративный процесс при БА, является теория β-амилоида [2]. Согласно данной концепции, цитотоксичные фрагменты β-амилоида длиной в 42 аминокислоты запускают в нейронах необратимый процесс самоагрегации тау-белка, в результате чего образуются нейрофибриллярные агрегаты, уменьшается плотность межсинаптических контактов и наступает атрофия серого вещества головного мозга. В свою очередь неоднократно предпринимались попытки объяснить сам процесс амилоидогенеза и развития тау-патологии, примером чего может служить теория митохондриального каскада [3]. Гипотеза первичного митохондриального каскада подразумевает, что образование β-амилоида и тау-протеина зависит от функциональной активности митохондрий клетки и ее биоэнергетического состояния, что, вероятно, связано с гидротопными свойствами АТФ [4]. Неудивительно, что изучению энергетического статуса нейронов и изменения митохондриальной функции постоянно уделяется много внимания. Однако совокупная митохондриальная функция может включать множество параметров, которые отражают различное состояние биоэнергетических процессов, активности систем детоксикации свободных радикалов и реакций апоптоза, происходящих в клетке. Учитывая трудоемкость экспериментальных подходов к моделированию БА, а также вариабельность показателей митохондриального каскада [5], выбор оптимального параметра позволит значительно упростить процедуры фармакологического скрининга активных соединений митохондриально-ориентированного действия. В связи с этим цель данного исследования состояла в оценке зависимости изменения показателей митохондриальной функции от различного содержания тау-белка в гиппокампе крыс в условиях экспериментальной БА спорадического типа.
Материал и методы
Работа выполнена на 40 самцах крыс Wistar массой тела 260–280 г, полученных из питомника лабораторных животных «Рапполово» (Россия). На время исследования животные содержались в стандартных условиях вивария в полипропиленовых боксах по 5 особей со свободным доступом к воде и корму при температуре окружающего воздуха 22±2 °С, относительной влажности 55–65% и 12‑часовом суточном цикле. Исследование проведено в соответствии с принципами ARRIVE 2.0. [6], содержание и проводимые с животными манипуляции — в соответствии с положениями, изложенными в Директиве 2010/63/EU Европейского парламента и Совета Европейского союза от 22 сентября 2010 г. по охране животных, используемых в научных целях. Животные были разделены на 2 равные группы по 20 особей: ЛО — ложнооперированные животные и БА — группа крыс, которым моделировали болезнь Альцгеймера. БА воспроизводили путем инъекции фрагментов β-амилоида (Аβ) в СА1‑часть гиппокампа (переднезадняя — 3,8 мм, медиально-латеральная — 2 мм, дорсально-вентральная — 2,6 мм от брегмы согласно G. Paxinos). Aβ1–42 (Sigma-Aldrich) растворяли в фосфатно-солевом буфере (pH 7,4) при температуре 4 °С. Полученный раствор непрерывно перемешивали в течение 36 ч до образования агрегатов. Животных наркотизировали хлоралгидратом (внутрибрюшинно, 350 мг/кг), скальпировали теменную область, проделывали трепанационное отверстие и помещали в стереотаксическую установку. Далее при помощи микродозатора с иглой G30 вводили Aβ1–42 в конечной концентрации 1 ммоль/л. Игла оставалась в месте инъекции в течение 5 мин, после чего ее извлекали. Рану ушивали и обрабатывали 10% раствором повидон-йода [7]. Во избежание дистресса во время операции животных помещали на согревающую подложку, после операции крысы находились под согревающей лампой. Оперативное вмешательство выполняли под общей анестезией, что сводило к минимуму возможность появления острого болевого синдрома.
Через 60 дней после операции у крыс оценивали когнитивные функции в тесте Y-образного лабиринта. Установка состояла из трех равных рукавов, соединенных под углом 120°. Животное помещали в центр установки и в течение 8 мин регистрировали число его перемещений между рукавами. При этом фиксировали спонтанные чередующиеся заходы в рукава (1-2-3, 3-1-2, 2-3-1). На основании полученных данных определяли процент спонтанного чередования (ПСЧ), который отражает изменение когнитивных способностей животных [8].

Далее крыс декапитировали под хлоралгидратной анестезией, извлекали головной мозг и выделяли гиппокамп. Гиппокамп гомогенизировали в среде, состоящей из 1 ммоль ЭДТА + 215 ммоль маннита + 75 ммоль сахарозы + 0,1% раствор БСА + 20 ммоль HEPES, рН 7,2. Полученный гомогенат разделяли на 2 части. Первую часть центрифугировали в режиме при 10 000 g в течение 10 мин и в полученном супернатанте определяли содержание тау-белка и АТФ. Вторую аликвоту гомогената использовали для получения митохондриальной фракции. Для выделения митохондрий гомогенат гиппокампа центрифугировали при 1100 g в течение 2 мин. Полученный супернатант в количестве 700 мкл переносили в пробирки Эппендорфа, смешивали с 75 мкл 10% перколла и центрифугировали при 18 000 g в течение 10 мин. Осадок ресуспендировали в 1 мл изолирующей среды и центрифугировали в течение 5 мин при 10 000 g. Все процедуры проводили при температуре 4 °C. [9] Во второй аликвоте оценивали изменение активности цитратсинтазы, аконитазы, сукцинатдегидрогеназы, цитохром‑с-оксидазы, концентрации пероксида водорода и митохондриального мембранного потенциала.
Активность аконитазы определяли спектрофометрически при 340 нм путем регистрации НАДФ·Н, образовавшегося в ходе сопряженной аконитаза-изоцитратдегидрогеназной реакции. Активность фермента рассчитывали по изменению оптической плотности, используя коэффициент экстинкции 0,0313 µМ–1 [10]. Активность цитратсинтазы оценивали спектрофотометрическим методом, основанным на определении окрашенных продуктов реакции деградации 5,5’-ди-тиобис-(2‑нитробензойной кислоты) в присутствии ацетил-КоА и оксалоацетата при 412 нм [11]. Активность цитохром‑с-оксидазы определяли в митохондриальной фракции по изменению оптической плотности среды реакции окисления цитохрома С (II) в присутствии KCN при 500 нм [12]. Активность сукцинатдегидрогеназы оценивали спектрофотометрически в реакции сукцинатзависимого восстановления дихлорфенолиндофенола при добавлении в анализируемую среду ротенона при 600 нм [13]. Концентрацию митохондриального пероксида водорода определяли методом флюоресцентного анализа с использованием стандартного набора Amplex Red (Thermo Fisher Scientific). Мембранный потенциал митохондрий оценивали спектрофотометрическим методом по изменению экстинкций проб с сафранином О при λ=515 нм и λ=525 нм [14]. Оптическую плотность образцов регистрировали на спектрофотометре Промэколаб ПЭ-5300В в кюветах с длиной оптического пути 10 мм.
Содержание АТФ и тау-белка оценивали методом твердофазного иммуноферментного анализа с применением видоспецифичных реактивов производства Cloud Clone (США). Регистрацию аналитического сигнала осуществляли с помощью микропланшетного ридера Infifinite F50 (Tecan, Австрия).
Полученные результаты обрабатывали методами вариационной статистики. В ходе статистического анализа использовали пакет прикладных программ STATISTICA 6.0. (StatSoft, США). Нормальность распределения данных оценивали с применением теста Шапиро–Уилка. Однородность дисперсий определяли, используя тест Левена. Статистически значимые отличия между группами оценивали методом однофакторного дисперсионного анализа с пост-тестом Ньюмена–Кейлса (при нормальном распределении данных) или пот-тестом Краскелла–Уоллиса (при распределении данных, отличных от нормального) при критическом уровне значимости р<0,05. Корреляционный анализ выполнен по Спирмену с интерпретацией значения коэффициентов корреляции по шкале Чеддока [15].
Результаты
В ходе работы было установлено, что у крыс с воспроизведенной БА в гиппокаме животных отмечается увеличение содержание тау-протеина в 7,5 раза (p<0,05) по сравнению с ЛО животными (табл. 1). Также в гиппокампе крыс с БА наблюдалось повышение содержания митохондриального пероксида водорода в 3,4 раза (p<0,05) по отношению к аналогичному показателю ЛО животных. В то же время в условиях экспериментальной БА в гиппокампе крыс отмечено статистически значимое уменьшение функциональной активности митохондрий, о чем свидетельствует отрицательная динамика изменения маркерных показателей. Так, активность цитратсинтазы у животных с БА снизилась на 88,5% (p<0,05) по сравнению с ЛО группой крыс, активность аконитазы, цитохром‑с-оксидазы и сукцинатдегидрогеназы также уменьшилась на 44% (p<0,05), 63,7% (p<0,05) и 67,8% (p<0,05) соответственно. Величина митохондриального мембранного потенциала и концентрация АТФ у крыс с экспериментальной БА были меньше аналогичных параметров ЛО животных на 71,3% (p<0,05) и 57,7% (p<0,05) соответственно. Стоит отметить, что у животных с БА через 60 дней после инъекции Аβ отмечено развитие когнитивного дефицита, о чем свидетельствует уменьшение (относительно ЛО крыс) количества спонтанного чередования (см. табл. 1) отсеков Y-образного лабиринта на 61,3% (p<0,05).
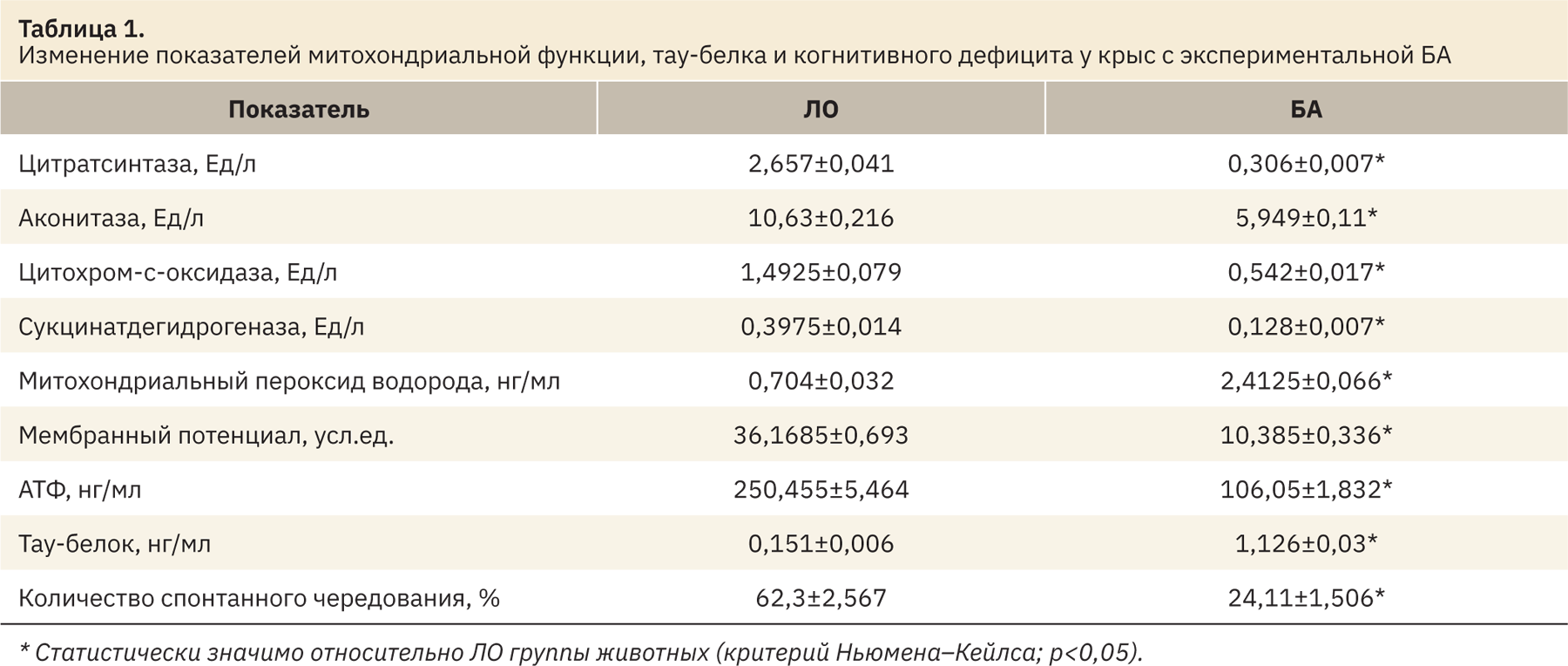
Проведенный корреляционный анализ позволил выявить следующие закономерности. Изменение когнитивного дефицита у крыс с БА очень сильно коррелирует с меняющейся концентрацией тау-белка в гиппокампе животных со значением r=0,9950 (рис. 1).
В свою очередь изменение концентрации тау-протеина очень сильно коррелирует с различной активностью цитратсинтазы и содержанием АТФ (рис. 2) в гиппокампе крыс (r=0,95806 и r=0,9798 соответственно). Также отмечена слабая корреляционная зависимость меняющейся концентрации тау-белка (табл. 2) от изменения активности цитохром‑с-оксидазы (r=0,346714) и концентрации митохондриального пероксида водорода (r=0,3907).
Обсуждение
Изменение реакций митохондриального каскада при БА является перспективным подходом к сохранению структурно-функциональной целостности нейронов. В то же время совокупная митохондриальная функция характеризуется множеством параметров, позволяющих оценить энергосинтетическую, редокс- и апоптозрегулирующую функцию митохондрий, что в ряде случаев может затруднять выбор целевого параметра при проведении фармакологических исследований. В связи с этим была выполнена данная работа, в которой исследовались корреляционные зависимости изменения показателей митохондриальной функции от концентрации тау-белка в гиппокампе крыс. В качестве экспериментальной модели БА выбрана модель спорадической БА, вызванной интрагиппокампальным введением Аβ. Данный вариант экспериментальной патологии использован как подход, позволяющий воспроизвести тау-патологию, выражающуюся в накоплении в нейронах нейрофибриллярных агрегатов гиперфосфорилированного тау-белка, что наиболее фенотипически близко к течению БА у людей [16]. В качестве митохондриальных маркеров были выбраны следующие: изменение концентрации АТФ как интегральный показатель, характеризующий функциональную активность митохондрий; активность цитратсинтазы — фермента-маркера синтеза митохондрий de novo; активность сукцинатдегидрогеназы и цитохром‑с-оксидазы — ферментативных маркеров митохондриального биогенеза и процессов митофагии; изменение активности аконитазы и концентрации митохондриального пероксида водорода, отражающее способность митохондрий регулировать окислительно-восстановительные процессы в клетке, а также изменение величины митохондриального мембранного потенциала как триггера митохондриально-зависимого пути апоптоза [17]. В итоге установлено, что у крыс со спорадической БА через 60 дней после введения Аβ отмечается развитие когнитивного дефицита, который очень сильно коррелирует с повышением концентрации тау-белка в гиппокампе крыс. В свою очередь изменение содержания тау-протеина очень сильно коррелирует с меняющейся активностью цитратсинтазы и концентрацией АТФ, в то время как для остальных изучаемых показателей установлена слабая корреляционная зависимость или отсутствие таковой. Данный факт может быть связан с тем, что активность цитратсинтазы и концентрация АТФ являются более интегральными показателями, чем остальные оцениваемые параметры, и отражают изменение совокупной митохондриальной функции [18], тогда как оставшиеся параметры практически изолированно характеризуют специфичные проявления митохондриальной дисфункции при БА: энергодефицит, окислительный стресс и повышение интенсивности реакций апоптоза.
Заключение
На основании полученных результатов можно предполагать, что в условиях экспериментальной БА спорадического фенотипа для оценки изменения митохондриальной дисфункции на ранних этапах целесообразно определять изменение активности цитратсинтазы и концентрации АТФ как показателей, наиболее сильно коррелирующих с различным содержанием тау-белка.
Список источников
- Scheltens P., De Strooper B., Kivipelto M. et al. Alzheimer’s disease // Lancet. 2021. Vol. 397. N. 10284. P. 1577–1590. DOI:10.1016/s0140-6736(20)32205-4.
- Lei P., Ayton S., Bush A.I. The essential elements of Alzheimer’s disease // J. Biol. Chem. 2021. Vol. 296. P. 100105. DOI: 10.1074/jbc.REV120.008207.
- Swerdlow R.H. Mitochondria and Mitochondrial Cascades in Alzheimer’s Disease // J. Alzheimers Dis. 2018. Vol. 62. N. 3. P. 1403–1416. DOI: 10.3233/JAD-170585.
- Rice A.M., Rosen M.K. ATP controls the crowd // Science. 2017. Vol. 356. N. 6339. P. 701–702. DOI: 10.1126/science.aan4223.
- Esquerda-Canals G., Montoliu-Gaya L., Güell-Bosch J., Villegas S. Mouse Models of Alzheimer’s Disease // J. Alzheimers Dis. 2017. Vol. 57. N. 4. P. 1171–1183. DOI: 10.3233/JAD-170045.
- Percie du Sert N., Hurst V., Ahluwalia A. et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research // PLoS Biol. 2020. Vol. 18. N. 7. P. e3000410. DOI: 10.1371/journal.pbio.3000410.
- Manczak M., Reddy P.H. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimer’s disease neurons: implications for mitochondrial dysfunction and neuronal damage // Hum. Mol. Genet. 2012. Vol. 21. N. 11. P. 2538–2547. DOI: 10.1093/hmg/dds072.
- Amani M., Zolghadrnasab M., Salari A.A. NMDA receptor in the hippocampus alters neurobehavioral phenotypes through inflammatory cytokines in rats with sporadic Alzheimer-like disease // Physiol. Behav. 2019. Vol. 202. P. 52–61. DOI: 10.1016/j.physbeh.2019.01.005.
- Connolly N.MC, Theurey P., Adam-Vizi V. Guidelines on experimental methods to assess mitochondrial dysfunction in cellular models of neurodegenerative diseases // Cell Death Differ. 2018. Vol. 25. N. 3. P. 542–572. DOI: 10.1038/s41418‑017‑0020‑4.
- Ternette N., Yang M., Laroyia M. Inhibition of mitochondrial aconitase by succination in fumarate hydratase deficiency // Cell Rep. 2013. Vol. 3. N. 3. P. 689–700. DOI: 10.1016/j.celrep.2013.02.013.
- Shepherd D., Garland P.B. The kinetic properties of citrate synthase from rat liver mitochondria // Biochem. J. 1969. Vol. 114. N. 3. P. 597–610. DOI: 10.1042/bj1140597.
- Li Y., D’Aurelio M., Deng J.H. An assembled complex IV maintains the stability and activity of complex I in mammalian mitochondria // J. Biol. Chem. 2007. Vol. 282. N. 24. P. 17557–17562. DOI: 10.1074/jbc.M701056200.
- Wang H., Huwaimel B., Verma K. Synthesis and Antineoplastic Evaluation of Mitochondrial Complex II (Succinate Dehydrogenase) Inhibitors Derived from Atpenin A5 // ChemMedChem. 2017. Vol. 12. N. 13. P. 1033–1044. DOI: 10.1002/cmdc.201700196.
- Zhyliuk V.I., Mamchur V.V., Pavlov S. Role of functional state of neuronal mitochondria of cerebral cortex in mechanisms of nootropic activity of neuroprotectors in rats with alloxan hyperglycemia // Eksp. i klin. farm. 2015. Vol. 78. P. 4–10.
- Akoglu H. User’s guide to correlation coefficients // Turk J. Emerg. Med. 2018. Vol. 18. N. 3. P. 91–93. DOI: 10.1016/j.tjem.2018.08.001.
- Pereira J.B., Janelidze S., Ossenkoppele R. et al. Untangling the association of amyloid-β and tau with synaptic and axonal loss in Alzheimer’s disease // Brain. 2021. Vol. 144. N. 1. P. 310–324. DOI: 10.1093/brain/awaa395.
- Molnar M.J., Kovacs G.G. Mitochondrial diseases // Handb Clin. Neurol. 2017. Vol. 145. P. 147–155. DOI: 10.1016/B978‑0‑12‑802395‑2.00010-9.
- Ranjbarvaziri S., Kooiker K.B., Ellenberger M. et al. Altered Cardiac Energetics and Mitochondrial Dysfunction in Hypertrophic Cardiomyopathy // Circulation. 2021. Vol. 144. N. 21. P. 1714–1731. DOI: 10.1161/CIRCULATIONAHA.121.053575
Поиск



