
Оценка мутагенности в тесте Эймса производного бензопентатиепина
Быков В.В., Седых И.М., Быкова А.В. Оценка мутагенности в тесте Эймса производного бензопентатиепина. Лабораторные животные для научных исследований. 2018; 4. https://doi.org/10.29296/2618723X-2018-04-01
Резюме
Резюме. Изучение мутагенных свойств нового лекарственного средства (ЛС) – важная часть доклинического исследования, поскольку предусматривает оценку способности ЛС к индукции разных типов мутаций в зародышевых и соматических клетках. При исследовании генотоксичности нового вещества особое внимание должно быть обращено на его структурное сходство с известными мутагенами. Для анализа структурного сходства используются, представительная база данных о мутагенных свойствах широкого круга химических соединений, а также специальные компьютерные программы, которые позволяют оценить взаимодействие исследуемого соединения или его метаболита с мишенями, обусловливающими мутагенный эффект. Экспериментальная оценка мутагенности кандидатов ЛС обязательно должна включать батарею минимум из 2 тестов, в том числе тест на индукцию хромосомных повреждений in vivo и тест на индукцию генных мутаций. В качестве последнего, как правило, служит тест Эймса, который является альтернативой использования лабораторных животных. С одной стороны, замена животных на менее организованные организмы является одним из принципов 3 Rs, с другой – использование бактериальных штаммов позволяет проводить тестирование в виде скрининга и в короткие сроки оценить потенциальную генотоксичность соединения. На основании этих данных может быть разработан дизайн дальнейших исследований генотоксичности и ряда других токсикологических тестов. В ходе работы оценивали способность тестируемого вещества к индукции мутаций у штаммов S. typhimurium (ТА98, ТА100, ТА1535, ТА1537) и E. сoli (WP2 uvrA и WP2 /pKM101) в 6 концентрациях: 15,6; 31,2; 62,4; 125,0; 250,0 и 500,0 мкг/мл. Для оценки потенциальной генотоксичности исследуемого вещества и его возможных метаболитов тестирование осуществляли в том числе и в условиях с метаболической активацией в присутствии S9 фракции печени крыс (индуктор Aroclor-1254) и без метаболической активации. По результатам исследования выявлено отсутствие мутагенных свойств на всех использованных тест-штаммах, однако не исключалось генотоксического действия при многократном применении.
Введение
Под мутагенностью веществ понимают их способность вызывать повреждение клетки и ее генетического материала, что проявляется изменением генотипа.
Существует большое количество веществ, обладающих мутагенной активностью, которые вносят структурные изменения
в хромосомы посредством
ДНК-реактивных или не-ДНК-реактивных механизмов. ДНК-реактивные процессы
увеличивают частоту хромосомных аберраций путем формирования ДНК-аддуктов через ковалентную связь непосредственно
с молекулой ДНК, в то время, как не-ДНК-реактивные механизмы вызывают повреждения ДНК путем таких
косвенных процессов, как, например, ингибирование ДНК-полимеразы, ингибирование топоизомеразы II, которое приводит
к двухцепочечным разрывам, или запуск перекисного окисления липидов [1].
Как известно, некоторые химические вещества, включая ксенобиотики, способны вызывать мутации в половых и соматических клетках человека. И если мутации в половых клетках повышают вероятность и частоту генетических или наследственных заболеваний, то наличие факта соматических мутаций дает основание предполагать канцерогенный потенциал исследуемого вещества. Реализовать свой мутагенный потенциал ксенобиотики могут через механизм замены пар оснований или непосредственной атаки (мутации могут происходить или в сайте исходной мутации, или в другом сайте хромосомы), а также через сдвиг рамки считывания (вставка или делеция одной или нескольких пар оснований в молекуле ДНК) [2].
Необходимо отметить, что хромосомные аберрации, включающие делеции (удаление участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую хромосому) потенциально могут приводить к канцерогенности [3]. Однако механизм канцерогенеза связан с хромосомной аберрацией, которая с вероятностью должна возникнуть в определенном месте, как например, в онкогене или опухолевом супрессоре. Таким образом, генотоксичность вещества далеко не всегда является маркером его канцерогенности. Кроме того, поскольку используемые при оценке генотоксичности in vitro бактерии являются прокариотами, это не идеальная модель для экстраполяции данных на млекопитающих, в том числе на человека [4]. Более того, использование в тесте Эймса бактерий с фенотипом rfa- (неполный синтез ЛПС) обеспечивает повышенную проницаемость бактериальной стенки для тестируемого вещества, что способствует к значительному повышению экспозиции вещества внутри клетки, в которой также созданы все условия для максимальной чувствительности к мутагенезу. Соответственно, положительный результат в тесте Эймса указывает лишь на генотоксичный эффект вещества in situ в идеальных условиях, и его необходимо трактовать лишь с позиций соответствия экспериментальных данных in vitro результатам анализа структурного сходства кандидата лекарственное средство (ЛС) с известными мутагенами. Можно предположить, что известное отсутствие мутагенности у млекопитающих, в том числе у человека, ряда генотоксичных in vitro ЛС [4] отражает различие биологических объектов и особенности фармакокинетики.
На наш взгляд, среди известных лабораторных воплощений теста Эймса для исследования индукции генных мутаций несомненным преимуществом обладает микропланшетный вариант. В стандартных наборах для его проведения используются качественные и сертифицированные штаммы микроорганизмов, а также малые количества исследуемого вещества, что важно на начальных этапах разработки инновационного лекарственного препарата, когда наработка испытуемых соединений проводится в лабораторных условиях. Кроме того, стандартные наборы могут применяться для автоматизированных тестеров, когда выполняются большие скрининговые программы [5].
Использование бактерий в качестве тест-систем – предмет интенсивного изучения многих лет [6].
Несмотря на кажущуюся простоту, бактерии, как
и эукариотические организмы, обладают сложным механизмом
реагирования на повреждение ДНК различными агентами [7]. Широкое распространение бактерии как тест-система получили
для исследования основных механизмов повреждения ДНК и как удобный скрининговый тест для исследования
потенциальных мутагенов [2].
Бактериальные тест-системы имеют ряд специфических характеристик, обусловливающих их чувствительность к мутагенам. Большинство канцерогенных соединений химически инертны per se [3, 4, 6]. Они часто метаболизируются до электрофильных промежуточных соединений, которые могут реагировать с такими клеточными макромолекулами, как белки, РНК и ДНК [7]. Однако указанная тест-система не в полной мере имитирует метаболизм исследуемого ксенобиотика, который протекает в клетках млекопитающих [8], в связи с чем появилась необходимость включения в инкубационную среду метаболической системы млекопитающих. Для этого используют постмитохондриальный супернатант (фракцию S9) печени крыс, обработанный предварительно комбинацией 3-метилхолантрена, фенобарбитала и β-нафтофлавона либо смесью полихлорированных бифенилов (например, Aroclor-1254), которые индуцируют пул необходимых метаболизирующих ферментов. Такой подход значительно увеличивает активность ферментов биотрансформации, вследствие чего повышается чувствительность к потенциальным мутагенам бактериальных тест-систем (рис. 1).
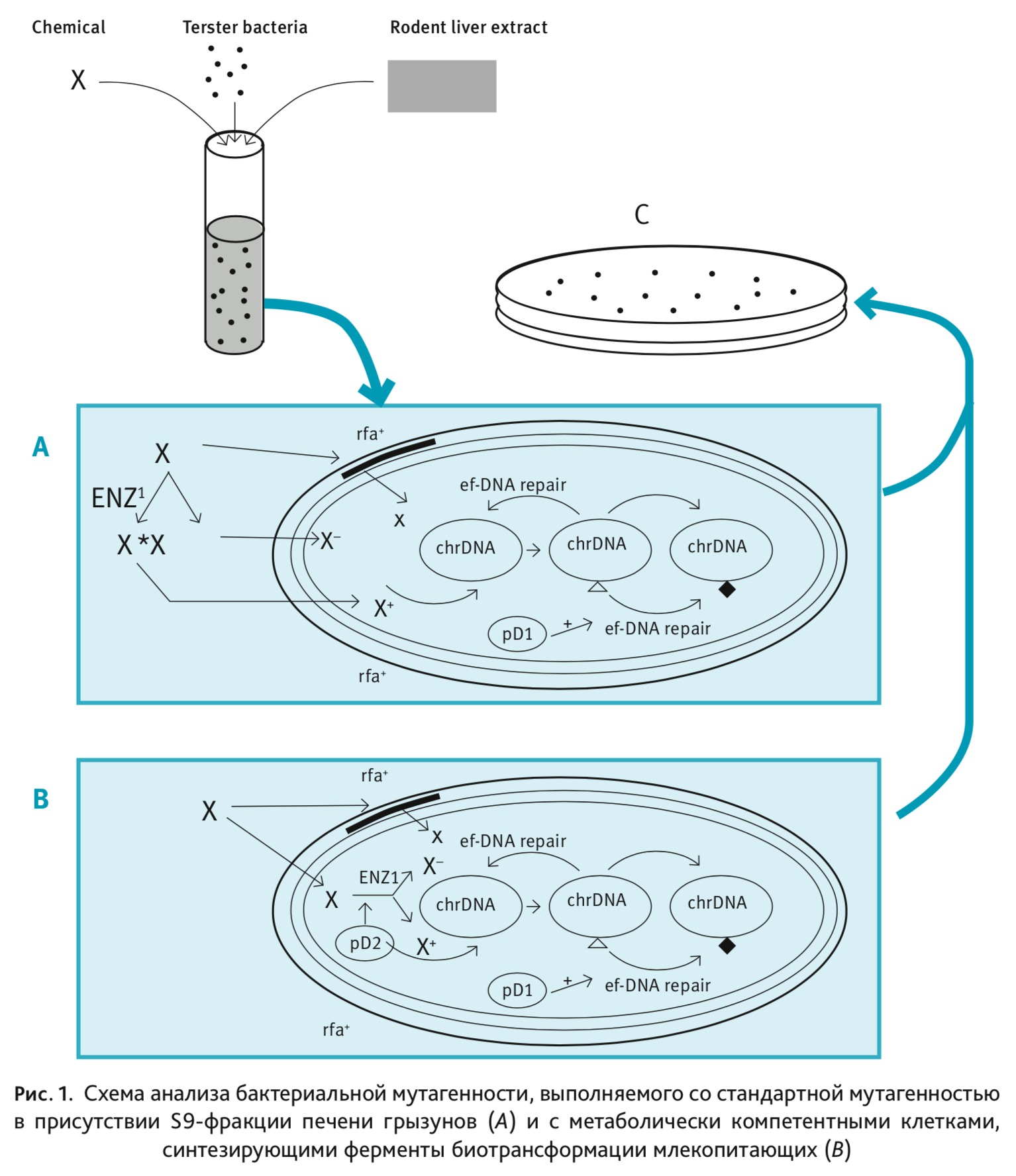
Наружная мембрана E. coli и S. typhimurium дикого типа препятствует проницаемости молекул
ксенобиотиков внутрь клетки (фенотип rfa+) [9, 10]. Мутации в опероне rfa (фенотип rfa-) приводят
к неполному синтезу клеточной мембраны (дефект синтеза липополисахаридной составляющей) и повышенной
проницаемости мембраны. Этот rfa-фенотип увеличивает чувствительность к мутагенам различного происхождения,
таким как ароматические амины, полициклические ароматические углеводороды и микотоксины, действие которых было
бы крайне сомнительно на клетки дикого ти-
па [11].
Активированный метаболит X* может вызвать повреждение (знак ∆) в хромосомной ДНК (chrDNA), результатом которого является мутация (знак ♦). Повреждение ДНК может быть устранено без ошибок (восстановление по пути ef-DNA), в результате чего ДНК восстановится до исходного состояния или процесс может пойти по пути «ремонта» (ep-DNA repair).
Большинство тест-бактерий содержат мутацию в одном из генов uvr, которая приводит к уменьшению резерва системы репарации ДНК, что также увеличивает их чувствительность к потенциальным мутагенам.
Бактерии также могут содержать плазмиду E. coli pKM101, которая увеличивает чувствительность бактерий к мутагенам и обеспечивает антибиотикорезистентность (в том числе к ампициллину, часто используемому в тесте Эймса).
В процессе тестирования ксенобиотиков на мутагенность чаще всего применяются бактерии с мутацией в генах биосинтеза аминокислот – бактерии с ауксотрофностью (trp, his или arg). Мутагенный эффект выражается в виде реверсии ауксотрофности для аминокислоты за счет усиления функции пораженного гена [12]. Различные целевые чувствительные аллели используются для конкретных мутаций, например, аллель hisG46 – для штамма TA100 S. typhimurium, аллель hisD 3052 – для штамма TA98 и т.д. Из-за специфики реализации и развития мутагенного эффекта возникает необходимость тестирования соединения на нескольких штаммах.
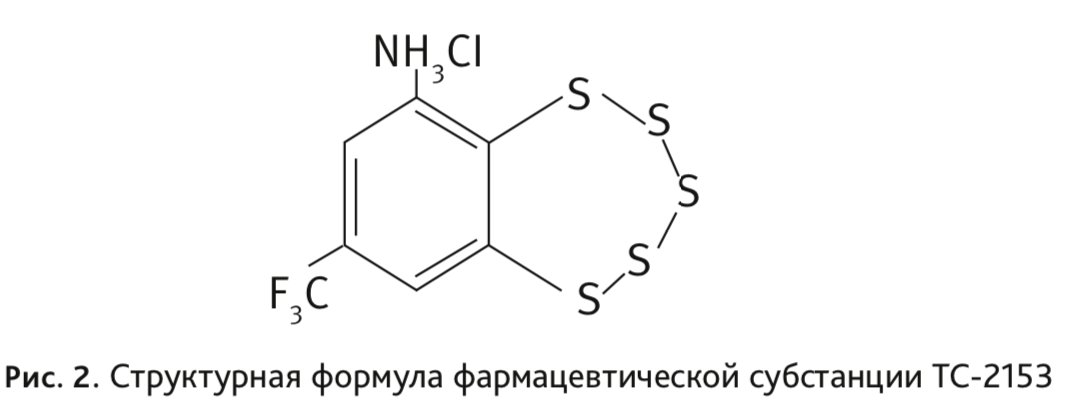
Тестированию на мутагенность подвергалась фармацевтическая субстанция TC-2153 – 8-(трифторметил) бензо[f] [1–5]пентатиепин-6-амингидрохлорид [13, 14] (рис. 2), которая исследуется нами при разработке лекарственного препарата для лечения депрессий, и любезно предоставлена для выполнения работы Отделом медицинской химии Новосибирского института органической химии им. Н.Н. Ворожцова СО РАН.
Соединение синтезировано по аналогии с цитотоксичным варацином (рис. 3), выделенным из морского асцидиана [15].
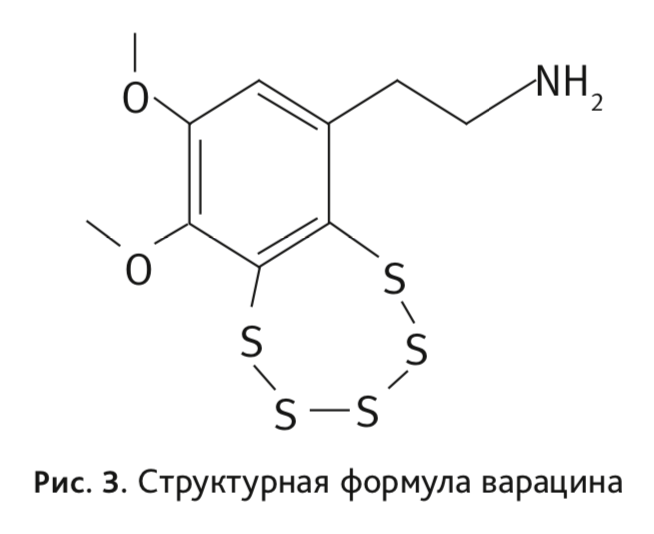
Цитотоксичность варацина связана с повреждением ДНК [16]. Уникальная пентатиепиновая часть молекулы варацина, присутствующая и в молекуле ТС-2153, обусловливает повреждение ДНК полисульфидным ионом [17] (рис. 4):
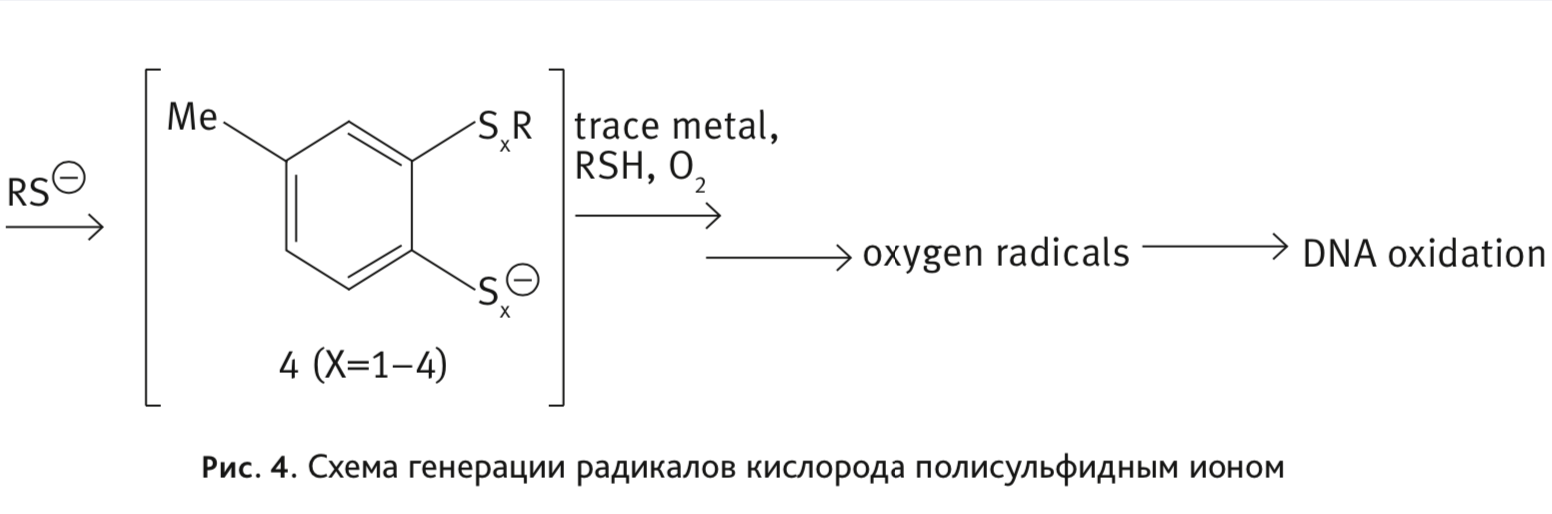
Предложенная схема повреждения ДНК, вызванная тиолом, связана с превращением молекулярного кислорода в реактивные радикалы с участием ионов Fe2+ в ходе реакции Фентона.
Согласно имеющимся данным, полимерный участок серы молекулы ТС-2153 может обладать потенциальной генотоксичностью не за счет прямого воздействия на ДНК, а посредством активации процесса перекисного окисления липидов.
Еще одним потенциально опасным с точки зрения возможной мутагенности участком молекулы TC-2153 является аминогруппа бензольного кольца. Известно, что ароматические амины (м-фенилендиамин, п-фенилендиамин; рис. 5) активируются до мутагенных метаболитов за счет гидроксилирования, идущего по ароматическому кольцу или по аминогруппе, а также за счет нитрования (наиболее известен прямой мутаген 4-нитро фенилендиамин) [18].
Для оценки возможной мутагенности ТС-2153 за счет гидроксилирования аминогруппы использовали прием добавки в инкубационную среду ферментов S9 фракции печени крыс [8].
Активированные мутагены (канцерогены) могут детоксицироваться, превращаясь в эфиры глюкуроновой кислоты (например, через гидроксильную группу после ее присоединения к бензольному кольцу в процессе взаимодействия с цитохромом P450) и выделяться почками [4]. Но возможное высвобождение активного мутагенного (канцерогенного) метаболита в водной среде (например, моче) за счет активности глюкуронидазы создает предпосылки развития онкологических патологий, в частности рака мочевого пузыря.
Сложное и зачастую непрямое взаимодействие ксенобиотика с системами биотрансформации, поддержания целостности генома, транскрипции и трансляции не позволяет, несмотря на значительный прогресс молекулярной биологии in silico, надежно прогнозировать канцерогенность разрабатываемого лекарственного препарата. Пожалуй, лишь сопоставление всего объема данных о новом соединении – химической структуре, тестах in vitro, in vivo, данных клинических исследований дает основание для принятия решения о возможности применения инновационного лекарства в клинической практике.
Материал и методы
В исследовании использовали коммерческий набор для проведения теста Эймса в микропланшетном формате (Ames MPF™ Penta I, Xenometrix, Швейцария). Исследование выполнено в соответствии с принципами GLP [19, 20].
В качестве тест-штаммов были выбраны 4 штамма S. typhimurium (ТА98, ТА100, ТА1535, ТА1537) и 1 штамм E. сoli (WP2 uvrA и WP2 /pKM101). Штаммы ТА98 (есть плазмида pKM101) и ТА1537 (нет плазмиды pKM101) подвержены мутации со сдвигом рамки считывания, штаммы ТА100, pKM101 (есть плазмида pKM101) и ТА1535, uvrA (нет плазмиды pKM101) подвержены мутации с заменой пар основания (мишень у сальмонелл GGG, а у кишечной палочки АТ). В то же время штаммы S. Typhimurium ауксотрофны по гистидину, в то время как E. coli – по триптофану.
Параллельное использование вышеназванных штаммов позволяет получить максимально полную информацию не только о факте генотоксичности молекулы, но и о характере этой генотоксичности.
Перед проведением тестирования на мутагенность получали стабильный раствор ТС-2153 с максимально возможной концентрацией, который не давал преципитации. После этого готовили линию разведений из ~10 концентраций, каждую из которых впоследствии тестировали на цитотоксичность в режимах –S9/(+S9) (для подбора максимальной концентрации, в которой не происходит гибели тест-штаммов от самой субстанции и/или ее возможных метаболитов). Максимальная концентрация, не обладающая цитотоксичностью, была стартовой в исследовании мутагенности. Следует отметить, что исследование мутагенности в тесте Эймса на высоких концентрациях, создание которых внутри клетки in vivo весьма сомнительно, по большому счету малоинформативно. Поэтому к старту исследования мутагенности желательно иметь данные фармакокинетики (экспозиции в крови, органах и распределения кровь/форменные элементы) для подбора необходимых тест-концентраций.
Субстанцию ТС-2153 на мутагенность тестировали в 6 концентрациях: 15,6; 31,2; 62,4; 125,0; 250,0 и 500,0 мкг/мл. Для каждого тест-штамма использовался свой позитивный контроль: 2-нитрофлуорен (2-NF), 4-нитрохинолин-N-оксид (4-NQO), N4-аминоцитидин (N4-ACT), 9-аминоакридин (9-AAc) и 2-аминоантрацен (2-АА).
Перед началом эксперимента оценивали приемлемость подготовленной ночной культуры тест-штаммов (наращивание ночной культуры проводили в течение не более 24 ч).
Оценку мутагенности осуществляли в присутствии S9 фракции печени крыс (для обнаружения мутагенности возможных метаболитов субстанции) и в ее отсутствии. В среду с S9 фракцией добавляли НАДФН-генерирующую систему (0,04 М НАДФ, 0,2 М глюкозо-6-фосфат, 0,25 М MgCl2), реакция происходила в среде 1 М KCl; и 0,2 М NaH2PO4.
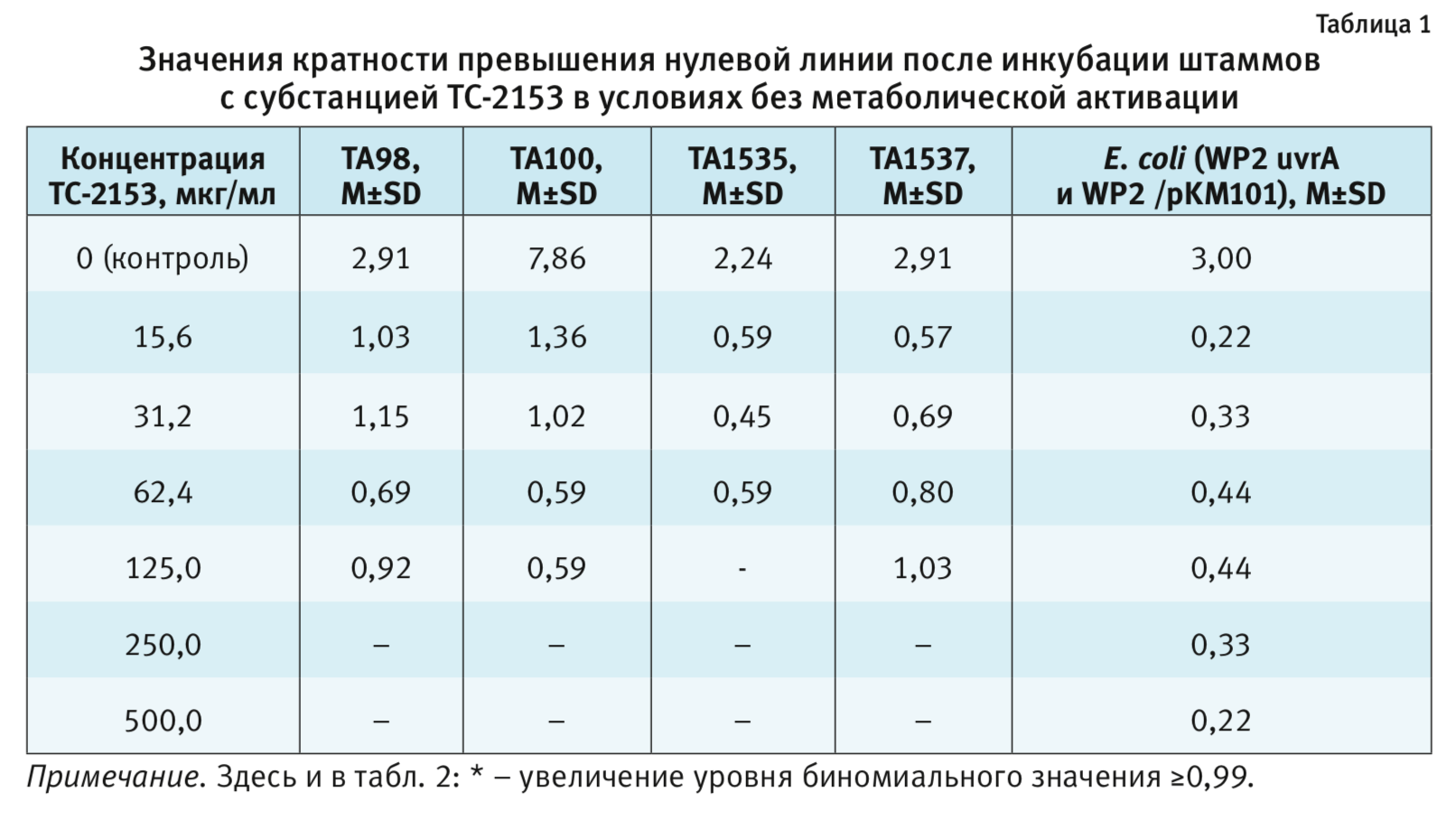
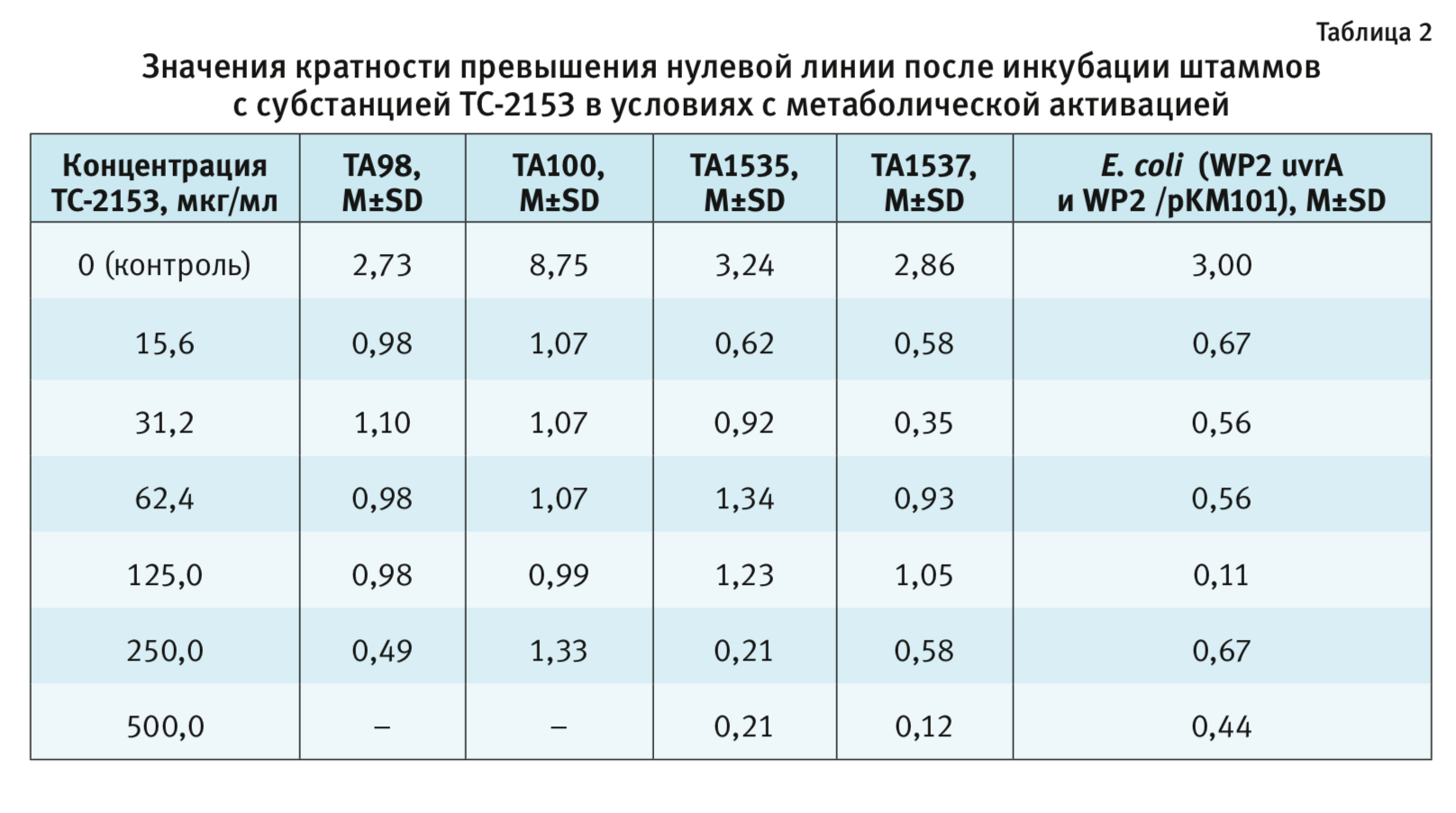
Критерием позитивного результата (наличия мутагенности) является зависимое от дозы превышение нулевой линии более чем в 2 раза при значении биномиального критерия не менее 0,99 (для исключения возможного спонтанного увеличения количества ревертантов) [21]
Результаты и обсуждение
При предварительном тестировании цитотоксичности ТC-2153 в условиях без метаболической активации было выявлено цитотоксическое действие в отношении штамма S. typhimurium ТА1535 в концентрациях 125,0; 250,0 и 500,0 мкг/мл, в отношении других штаммов S. typhimurium (ТА98, ТА100, ТА1537) – в концентрациях 250,0 и 500,0 мкг/мл; поэтому мутагенность в этих концентрациях не исследовалась.
В условиях без метаболической активации (табл. 1) показано, что субстанция ни в одной из концентраций не
вызывает увеличения количества ревертантов штаммов S. typhimurium ТА98, ТА100, TA1535, ТА1537 и E.
coli (WP2 uvrA и
WP2/pKM101) относительно негативного контроля.
При предварительном тестировании цитотоксичности TC-2153 в условиях с метаболической активацией выявлено
цитотоксическое действие субстанции
в отношении штаммов S. typhimurium ТА98
и ТА100 в концентрации 500 мкг/мл,
поэтому мутагенность в этой концентрации не
исследовалась.
В условиях с метаболической активацией (табл. 2) показано, что субстанция ни в одной
из концентраций не вызывает увеличения количества ревертантов штаммов S. typhimurium ТА98, ТА100, TA1535,
ТА1537 и E. coli (WP2 uvrA и
WP2/pKM101) относительно негативного контроля.
Результаты исследования указывают на отсутствие мутагенности субстанции TC-2153 в отношении использованных тест-штаммов. Такие данные говорят об отсутствии трансформации аминогруппы бензольного кольца в гидроксипроизводное и генерации радикалов кислорода полисульфидным ионом. Потенциальное окисление бензольного кольца до образования пирокатехин-производных соединений (сильные мутагены), по-видимому, идет в недостаточной степени для реализации генотоксического эффекта.
Возможно, что нуклеофильные атаки по бензольному кольцу в орто- и параположении относительно аминогруппы затруднены наличием крупных соседних заместителей (пентатиепиновым и относительно инертным трифторметильным), что также затрудняет образование генотоксичных метаболитов.
На основании полученных данных можно предположить, что однократный прием ТС-2153 даже в высокой дозе
(обеспечение экспозиции в организме выше
15 мкг/мл довольно затруднительно) не будет вызывать
генотоксического эффекта. Однако нельзя утверждать, что он не проявится при длительном применении исследуемого
соединения в терапевтической дозе, поскольку пентатиепиновый участок молекулы достаточно лабилен и его
отрыв (частичный или полный) может снять стериоконформационную затрудненность с молекулы и ускорить (или
запустить) процесс метаболизма с наработкой указанных выше метаболитов. Если это произойдет и данные
соединения будут обладать кумуляцией, а механизмы репарации ДНК и сдерживания перекисного окисления не
справятся с предотвращением развития генотоксичности, то в хроническом эксперименте либо при длительном
приеме пациентом не исключен мутагенный эффект. Однако более обоснованно судить о мутагенности ТС-2153 можно
только после оценки в принятых тестах репродуктивной токсичности и канцерогенности. Но даже в случае
положительных результатов последних тестов прямая экстраполяция их на человека невозможна. Различия в системах
биотрансформации и защиты ДНК от прямых повреждений и действия свободных радикалов требуют рассмотрения
генотоксичности и канцерогенности лекарства лишь по совокупной оценке данных in silico, in vitro, in vivo,
в клинических исследованиях, а также по результатам фармаконадзора за применяемым в медицинской
практике препаратом.
Однако даже наличие мутагенного эффекта в указанном нами тесте не должно восприниматься как объективно подтвержденный мутагенный (потенциально канцерогенный) эффект лекарственного кандидата и предпосылкой к приостановке или окончанию его разработки. На данный момент на рынке имеется широкий спектр препаратов с подтвержденной генотоксичностью, причем не относящихся к препаратам для лечения онкозаболеваний [22–24], которыми люди успешно лечатся десятки лет. Такое положение вещей указывает на то, что для проявления потенциальной генотоксичности и канцерогенности у человека необходимо создание особых условий – высокая системная экспозиция (часто неадекватно терапевтическим дозам), снижение функционального резерва защитных ферментативных систем, чрезмерно долгие курсы приема препарата (выше рекомендуемых), а также чрезмерная индукция ферментных систем детоксикации ксенобиотиков [25]. Совместное воздействие последних, несомненно, маловероятно.
Заключение
В ходе анализа структуры молекулы фармацевтической субстанции TC-2153 на предмет обнаружения маркеров потенциальной генотоксичности были выявлены опасные участки. В тесте Эймса в концентрациях 15,6–500,0 мкг/мл генотоксичность ТС-2153 не выявлена. Учитывая, что при ожидаемой терапевтической дозе ТС-2153 порядка 10,0 мг/кг для крысы концентрация вещества в крови и тканях при распределении в организме не достигнет 500,0 мкг на 1 мл крови или на 1 мг ткани, можно предположить отсутствие мутагенных эффектов ТС-2153 при терапевтическом применении. Однако, учитывая возможную избирательность накопления TC-2153 в тканях в ходе курсового лечения, для более обоснованного суждения о мутагенности субстанции ТС-2153 необходимо исследовать его влияние на репродуктивные свойства животных (генеративную токсичность и эмбриотоксичность), а также оценить канцерогенность in vivo.
Список источников
- Freeman B.A., Crapo J.D. Free radical and tissue injury. Adv. Biol. Disease. 1984; 1: 26–40.
- Barnes D.E., Lindahl T., Segwick B. DNA Repair. Curr. Opin. Cell Biol. 1993; 5: 424–33.
- Guengerich, F.P. Metabolic activation of carcinogens. Pharmac. Ther., 1992; 54: 17–61.
- Miller E.C. and Miller J.A. Searches for ultimate carcinogens and their reaction with cellular macromolecules. Cancer. 1981; 47: 2327–45.
- Руководство по проведению доклинических исследований лекарственных средств, ч. I.
- Zhengyin Yan, Gary Caldwell. Optimization in drug discovery: in vitro methods. Chapter «Improvement of the Ames test using human liver S9 preparation». Methods in pharmacology and toxicology. Humana Press, 2004.
- Venitt S., Bartsch H., Becking G. et. al. Short-term assays using bacteria. In: LongTerm and Short-Term Assays for Carcinogens: A Critical Appraisal. IARC Scientific Publications, no. 83, Lyon, Chap. 5, 1986. Oxford University Press, Oxford, UK: 143–56.
- Anderson, D. Genotoxicity assays. In: Molecular Aspects of Oxidative Drug Metabolizing Enzymes. Arinç, E., Schenkman, J.B., and Hodgson, E., Eds., NATO ASI Series, 1995; H90, Springer-Verlag, Berlin: 303–96.
- Austin E.A., Graves J.M., Hite L.A., Parker C.T., and Schnaitman C.A. Genetic analyses of lipopolysaccharide core biosynthesis by Escherichia coli K12: insertion mutagenesis of th rfa locus. J. Bacteriol. 1990; 172: 5312–25.
- Parker C.T., Kloser A.W., Schnaitman C.A., Stein M.A., Gottesman S., and Gibson B.W. Role of the rfaG and rfaP genes in determining the lipopolysaccharide core structure and cell surface propertie s of Escherichia coli K-12. 1992.
- Anderson D. Genotoxicity assays. In: Molecular Aspects of Oxidative Drug Metabolizing Enzymes.
- Arinc E., Schenkman J.B. and Hodgson E., Eds., NATO ASI Series. 1995; H90, Springer-Verlag, Berlin: 303–96.
- Maron D.M. and Ames B.N. Revised methods for Salmonella mutagenicity test. Mutat. Res. 1983; 113: 173–215.
- Khomenko T.N., Korchagina D.V., Komarova N.I., Volcho K.P., Salakhutdinov N.F. Synthesis of 6-Amino-benzopentathiepines by Reactions of 4-Nitro-benzyldithiol-2-ones with NaHS Lett. Org. Chem. 2011; 8 (3): 193–7.
- Kulikova E.A., Volcho K.P., Salakhutdinov N.F., Kulikov A.V. Benzopentathiepine Derivative, 8-(Trifluoromethyl)-1,2,3,4,5-Benzopentathiepin-6-Amine Hydrochloride (TC-2153), as a Promising Anti-depressant of New Generation Letters in Drug Design & Discovery. 2017; 14; 8: 974–84 DOI:10.2174/1570180814666161121112417.
- Davidson B.S.; Molinski T.F.; Barrows L.R.; Ireland C.M. J. Am. Chem. Soc. 1991; 113: 4709–10.
- Barrows L.R.; Paxton M.B.; Kennedy K.A.; Thompson L.H. Carcinogenesis. 1991; 12: 805.
- Sato R.; Ohyama T.; Ogawa S. Heterocycles 1995; 416 893–6.
- Ang E.L., Obbard J.P., Zhao H.M. Probing the molecular determinants of aniline dioxygenase substrate specificity by saturation mutagenesis. FEBS J. 2007; 274: 928–93.
- Гуськова Т.Д. Оценка безопасности лекарственных средств на стадии доклинического изучения. Химико-фармацевтич. Журн., 1990; 7: 10–5.
- OECD Bacterial Reverse Mutation Test 471, Adopted 21st July. 1997.
- Инструкция по применению теста Эймса в микропланшетном формате Ames MPF™ Penta I. S. typhimurium TA98, TA100, TA1535, TA1537, E. coli uvrA + E. coli /pKM101. Xenometrix. Версия 5.5, январь 2017.
- Edwards D.I. Reduction of nitroimidazoles and DNA damage. Biochem Pharmacol. 1986; 35: 53–8.
- Lopez Nigro M.M., Carballo M.A. Genotoxicity and cell death induced by tinidazole (TNZ). ToxicolLett. 2008; 180: 46–52.
- Alja Plosnik, Marjan Vracko, Marija Sollner Dolenc, Mutagenic and carcinogenic structural alerts and their mechanisms of action, Plosnik A, Vracko M, Sollner Dolenc M. Mutagenic and carcinogenic structural alerts and their mechanisms of action Arh Hig Rada Toksikol 2016; 67: 169–82.
- Кобляков В.А. Индукторы суперсемейства цитохрома p450 как промоторы канцерогенеза. Биохимия, 1998; 63 (8): 1043–58.
Поиск



