Колебания биохимических показателей при изменении гелиогеофизической обстановки
Павлова Р.Н., Мурзина А.А., Дадали В.А., Соколова Е.А. Колебания биохимических показателей при изменении гелиогеофизической обстановки. Лабораторные животные для научных исследований. 2020; 1. https://doi.org/10.29296/2618723X-2020-01-08
Резюме
Известно, что в процессе срочной и долговременной адаптации к факторам внешней среды, в первую очередь к гелиогеофизическим, наибольшим изменениям подвержены биохимические показатели, обеспечивающие гомеостаз. Одним из неучтенных факторов, влияющих на формирование ответной реакции биохимической системы (при стандартных условиях проведения эксперимента), является гелиофизическая обстановка. В дни резкого изменения гелиогеофизических параметров могут наблюдаться изменения величины биохимических показателей в контрольной группе и изменения характера ответной реакции биологической системы. Рассматриваются причины, вызывающие такие изменения в контрольной группе и ответную реакцию на воздействие в стандартных условиях эксперимента, приводится сравнение с предыдущими данными.
Представлен ретроспективный обзор собственных данных, полученных в период с 1975 по 2009 г. по колебаниям биохимических показателей при изменении гелиогеофизической обстановки. Рассмотрены данные, полученные в исследованиях ex vivo и in vivo; объектом исследования служил гомогенат мозга и гемолизат крови беспородных белых крыс. В ходе ретроспективного анализа было оценено влияние гелиогеофизической обстановки на активность супероксиддисмутазы гемолизата крови и общей аденозинтрифосфатазы мозга крыс, содержание небелковых SН-групп гемолизата крови. Проведен корреляционный анализ колебаний биохимических и гелиофизических показателей. В ходе исследования показано, что в периоды гелиофизических флуктуаций происходит не только изменение активности ферментов (супероксиддисмутазы гемолизата крови и аденозинтрифосфатазы головного мозга крыс), но и изменение характера ответной реакции на физическое или химическое воздействие – появление разнонаправленной ответной реакции на воздействие фактора при тех же (кроме гелиофизической обстановки) условиях эксперимента. Появление в отдельные дни в серии биохимического эксперимента атипичного ответа может быть обусловлено космофизическими воздействиями, что доказывает сопряженность биохимических параметров с гелиогеофизической средой.
Введение
Важнейшим условием при исследовании действия любого рода факторов физической или химической природы на организм человека и животных является определение показателей в контрольной группе у здоровых. В настоящее время введена строгая стандартизация по отбору и условиям содержания животных контрольной и экспериментальной групп [1]. Однако имеются факторы, не поддающиеся стандартизации и контролю, в частности космофизические воздействия. Многочисленные исследования показали наличие корреляции между увеличением числа госпитализированных больных с разными заболеваниями, в первую очередь с сердечно-сосудистыми и нервно-психическими расстройствами, и другими патологиями, и колебаниями гелиофизических показателей. Возможность ответа биологических систем на гелиогеофизические факторы подтверждена в многочисленных исследованиях, результаты которых опубликованы [2–4].
В дни резкого изменения гелиогеофизических параметров может наблюдаться изменение биохимических параметров в контрольной группе, выраженность ответа или смена знака ответной реакции биологической системы на изучаемое воздействие. Однако, в связи со сложностью экспериментальной реализации задачи, работ, демонстрирующих не только количественное изменение показателей в период гелиофизических флуктуаций, но и смену знака ответной реакции, в цитируемых периодических изданиях практически нет. В нашей лаборатории проводились исследования, в которых удалось обнаружить смену знака, т.е. изменение характера ответной реакции биологического объекта, в частности активности ряда ферментов при изучении воздействия на них ряда физических и химических факторов в период выраженных гелиофизических флуктуаций.
Цель работы – анализ причин, вызывающих изменение величин показателей в контрольной группе и изменение ответной реакции на воздействие в стандартных условиях эксперимента по сравнению с предыдущими данными.
В статье представлен ретроспективный обзор собственных данных, полученных в период с 1975 по 2009 г. по колебаниям биохимических показателей при изменении гелиогеофизической обстановки.
Результаты и обсуждение
При исследовании активности АТФазы мозга крыс нами обнаружены значительные колебания показателя в контрольной группе животных (табл. 1).
Таблица 1
Колебания активности АТФазы в гомогенатах мозга крыс и в эритроцитах барана в зависимости от гелиофизических флуктуаций
|
Месяц |
Активность общей АТФазы мозга крыс (мкг Р/мг белка•ч) |
Активность общей АТФазы эритроцитов барана (мкг Р/мг белка•ч) |
ВПОУ |
Число хромосферных вспышек |
|
Май |
39,5±4,0 |
– |
290±11 |
30 |
|
Июнь |
– |
31,5±4,2 |
316±26 |
25 |
|
Сентябрь |
10,7±3,8 |
10,9±9,7 |
118±27 |
80 |
|
Ноябрь |
26,7±13,9 |
– |
218±33 |
60 |
Примечание. ВПОУ – величина времени полуокисления унитиола.
В табл. 1 приведены резкие колебания активности общей АТФазы мозга крыс в разные месяцы 1975 г. Активность общей АТФазы нервной ткани определяли по скорости отщепления неорганического фосфора и выражали в мкг фосфора на мг белка в час. Активность АТФазы имеет хорошую корреляцию с числом хромосферных вспышек (ранговый коэффициент корреляции 0,91). Отметим, что изменения не носят сезонного характера так как в течение одного сезона (осень 1975 г.) активность фермента в сентябре и ноябре отличалась в 2,5 раза. В тот же период времени, Родионова Л.П. в другой лаборатории проводила определение активности АТФазы эритроцитов барана; колебания активности носили схожий характер [4]. Кроме того, приведены величины времени полуокисления унитиола – теста, характеризующего окислительно-восстановительное состояние среды, которое проявляется в конформационных изменениях биологических систем (см. табл. 1). Величина ВПОУ в тот период времени определялась ежедневно [2, 4].
Кроме изменений активности фермента в период гелиофизических флуктуаций в разные периоды 1975 г. имеет место изменение характера ответной реакции АТФазы на физическое воздействие – низкочастотное электромагнитное поле (ПеМП) частотой 0,5 гц, плотностью потока мощности 0,8.10-3 мвт /см3, напряженностью поля в зоне однородности 2,1э. Время экспозиции гомогената мозга крыс в ПеМП – 6 ч при температуре 20˚С (табл. 2).
Таблица 2
Изменение активности АТФазы мозга крыс в опытах ex vivo при действии низкочастотного электромагнитного поля (частота встречаемости эффекта)
|
Дата |
Активация, % |
Отсутствие эффекта, % |
Ингибирование, % |
|
Май, 1975 г. |
Нет |
10 |
90% |
|
Сентябрь, 1975 г. |
36 |
19 |
45 |
|
Ноябрь, 1975 г. |
18 |
37 |
45 |
Таким образом, нам удалось обнаружить, что в периоды гелиофизических флуктуаций происходит не только количественное изменение активности фермента, но меняется характер ответной реакции на физическое воздействие – появляется разнонаправленная ответная реакция на воздействие фактора при тех же (кроме гелиофизической обстановки) условиях эксперимента.
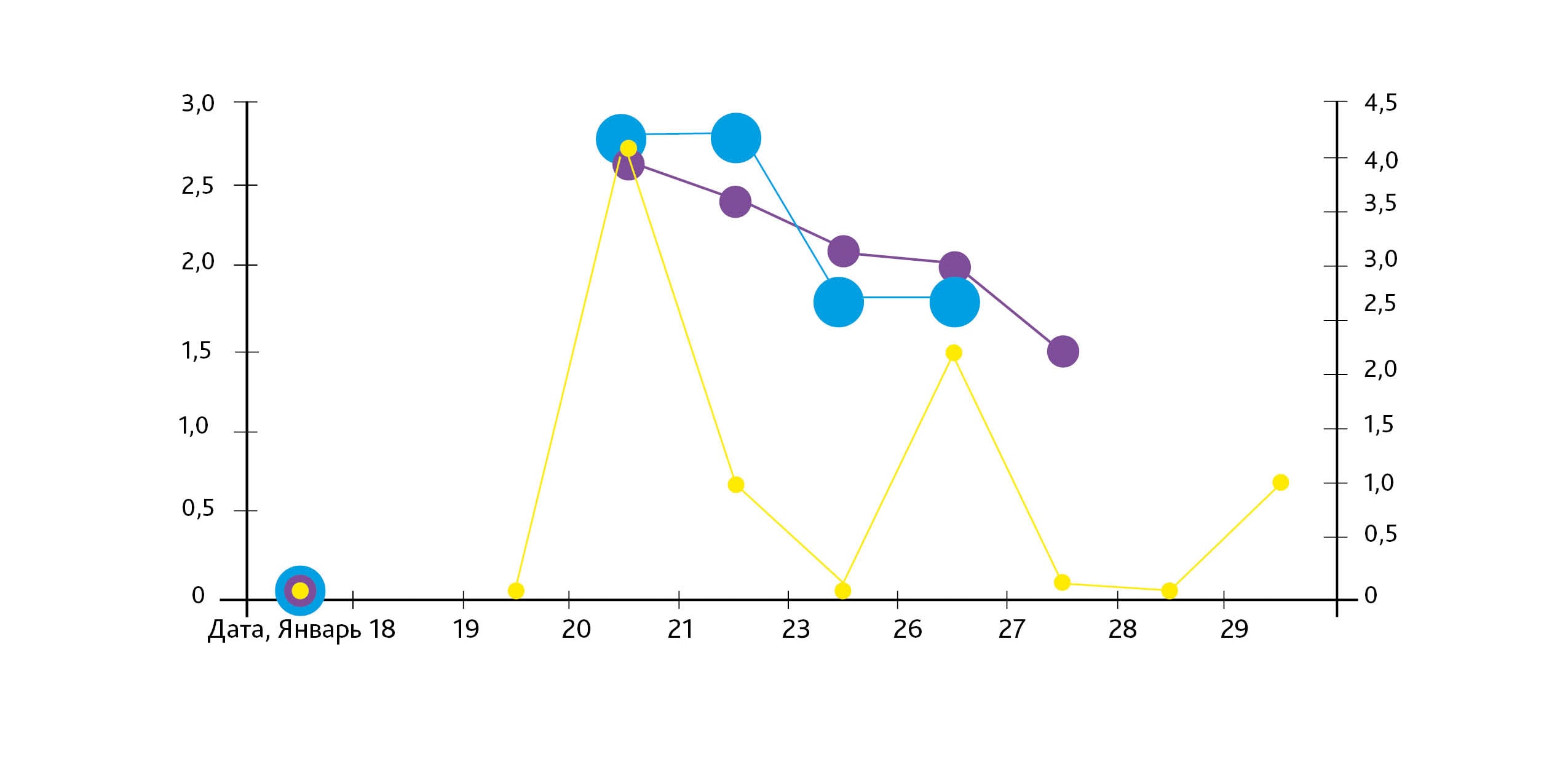
Другим примером изменения характера ответной реакции биологической системы на изучаемый фактор в период флуктуаций гелиогеофизических параметров является изменение знака ответа суперосиддисмутазы (СОД) и небелковых SH-групп крови крыс. В проведенных в январе 2009 г. опытах in vivo изучали изменения данных показателей на действие трансфер-фактора (пептидного комплекса с иммунотропными и адаптогенными свойствами), вводимого в дозе 0,07 мг/г интрагастрально в течение 10 дней до исследования [5]. При анализе результатов обнаружились выраженные колебания показателей антиоксидантной системы (АОС) у животных контрольной группы (ежедневно контрольная группа состояла из 3 животных) в период эксперимента: супероксиддисмутазы (СОД), каталазы, глюкозо-6-фосфатдегидрогеназы, глутатион-S-трансферазы и содержания белковых и небелковых SH-групп [6, 7]. Поэтому сопоставляли гелиофизические флуктуации с динамикой показателей АОС в период эксперимента. Активность СОД (у.е./мг белка) определяли определяли спектрофотометрически с квертицином [8], содержание SН-групп – методом амперометрического титрования [9].
Таблица 3
Активность СОД, концентрация небелковых SH-групп крови крыс при действии трансфер-фактора и величины чисел Вольфа и К-индекса магнитного поля Земли в январе 2009 г.
|
Показатель/дата |
19.01 |
20.01 |
21.01 |
23.01 |
26.01 |
27.01 |
28.01 |
|
СОД (у.е.) Контроль |
4,32±0,17 |
4,48±0,15 |
10,63*±3,57 |
9,37*±0,51 |
5,43±0,54 |
6,64±0,41 |
5,32±0,55 |
|
СОД (у.е.) Трансфер-фактор |
– |
2,36±0,06 |
5,45*±0,03 |
4,86±1,53 |
6,25*±0,12 |
– |
– |
|
Небелковый SH-гр. (мкм/мл) Контроль |
– |
2,60±0,03 |
2,40±0,02 |
2,10±0,03 |
2,0±0,03 |
1,5 |
– |
|
Небелк. SH-гр. Трансф. фактор |
– |
2,8±0,02 |
2,8±0,02 |
1,8*±0,034 |
1,8*±0,05 |
– |
– |
|
Число Вольфа |
13 |
0 |
0 |
0 |
0 |
0 |
0 |
|
К-индекс геомагнитного поля Земли |
41101322* |
10011011 |
00012211 |
0000000 |
22324210* |
01000221 |
10010101 |
Примечание. Представлены среднеарифметические величины и ± доверительный интервал; * – статистически значимые отличия при р0,05 (t-критерий Стьюдента).
В табл. 3 представлены средние величины показателей 3 животных, полученные в данный день, и величины гелиогеофизических параметров января 2009 г., цитированные по обзору состояния солнечной активности, ионосферы и магнитного поля Земли за январь 2009 г. данные предоставлены Новосибирским центром по гидрометеорологии и мониторингу окружающей среды для библиотеки Государственной геофизической обсерватории Санкт-Петербурга).
По этим данным 19.01.2009 имела место выраженная хромосферная вспышка. В предшествующую неделю величина чисел Вольфа была равна 0, величина К-индекса геомагнитного поля Земли колебалась в пределах 00000110–11210232. Согласно данным табл. 3, активность СОД в контроле через день после вспышки резко увеличилась, 21.01 и 23.01 трансфер-фактор снижал активность фермента, удерживая в пределах контроля предыдущих дней, однако 26.01 в ответ на действие трансфер-фактора в той же дозе произошла активация фермента.
Концентрация небелковых SH-групп гемолизата крови в контроле в период с 19.01.09 по 27.01.09 ежедневно снижалась, 20.01 и 21.01, действие трансфер-фактора приводило к повышению содержания SH-групп гемолизата крови, а 23.01 и 26.01 при тех же условиях эксперимента содержание небелковых SH-групп гемолизата крови в экспериментальной группе при действии трансфер-фактора снизилось по сравнению с контролем (см. рисунок).
Среди возможных механизмов формирования разнонаправленного ответа на действие факторов малой интенсивности важную роль играет исходное конформационное состояние молекулы белка, которое может меняться при действии ряда факторов и определять «поведение» белковой молекулы: изменения рН среды [10], изменения окислительно-восстановительного потенциала среды, в частности изменение тиолдисульфидного равновесия [2, 4], изменение липидного окружения мембранных ферментов [11], изменения гелиофизических параметров, в частности ряд биохимических показателей имеет четко выраженные суточные и сезонные колебания [12, 13].
В работе [14] показаны колебания активности АТФазы мышечной ткани, периодический ход которых обусловлен изменением концентраций конечных (или промежуточных) метаболитов, вызывающие конформационные перестройки в молекуле фермента, являющиеся причиной изменения ответной реакции.
Изменения рН среды, сопровождающиеся конформационными перестройками белковой молекулы, могут приводить к появлению иного типа ответа фермента, например ацетилхолинэстеразы на действие неорганических солей [10], или изменение ингибирования на активацию глутатион –S-трансферазы на действие солей желчных кислот [16].
Окислительно-восстановительный потенциал среды выполняет регуляторную функцию в клеточных процессах [2, 4]. Исследователи [15] установили, что при действии восстановителя имеет место повышение, а при действии окислителя – снижение активности АТФаз митохондрий мозга крыс. Очень часто не учитывают такой важный определяющий поведение мембранных белков фактор, как их липидное окружение [11].
При изменении гелиофизической ситуации в ряде случаев происходит окислительная модификация небелковых и белковых тиоловых групп, сопровождающаяся конформационными изменениями белков и ферментов и изменением их активности. Все эти факторы могут способствовать изменению характера ответной реакции биологического объекта на воздействие в условиях изменения гелиофизической обстановки и быть одной из причин, объясняющих наличие разного типа ответа при одинаковых воздействиях, что отмечалось разными авторами.
Вклад авторов: все авторы внесли существенный вклад в работу над статьей и несут ответственность за ее содержание.
Список источников
- Международный стандарт GLP “OECD Principles on Good Laboratory Practice” (OECD, ENV/MC/CHEM(98)17, 1997) США. Good Laboratory Practice for Nonclinical Laboratory Studies ( FDA, 21 CFR Part 58, 22 dec. 1978)
- Горшков Э.С. Редокс реакции в космобиологии / Э.С. Горшков, В.В. Иванов, В.В. Соколовский, СПб.: Издательство Политехнического университета, 2014 – 194 с.
- Жвирблис В.Е. О воспроизводимости гелиобиологических экспериментов. // Проблемы космической биологии. Л: Наука, 1989. - С. 145-160.
- Соколовский В.В. Тиолдисульфидная система в реакциях организма на факторы окружающей среды / В.В. Соколовский. - Санкт-Петербург.: Наука - 2008. - 112 с.
- Исследования адаптогенных и антиоксидантных свойств трансфер-фактора «Эдвенсд» / В.А Дадали, О.А. Агафонова, Н.Э. Голованова, Р.Н. Павлова, В.А. Кулеба, М.Н. Смертина, Ч.Р. Бейшебаева // Вопросы биологической, медицинской и фармацевтической химии. – 2011. - № 11 – С. 64-70.
- Коррегирующее действие трансфер-фактора «Эдвансд» в период гелиофизических флуктуаций / Р.Н. Павлова, О.А. Агафонова, Н.Э. Голованова, и др.// в сб. Современные подходы к метаболической коррекции в профилактике и терапии. – СПб:. – 2009 - С. 90-96.
- Вариации характера ответной реакции биологической системы при изменении космофизических факторов / Р.Н, Павлова, В.А. Дадали, В.А. Кулеба, Т.А. Махова, А.А. Мурзина //Труды У1 Международного конгресса «Слабые и сверхслабые поля и взаимодействия в биологии и медицине» СПб . - 2012. - С. 175.
- Арутюнян А.В. Методы оценки свободнорадикального окисления и антиоксидантной системы / А.В. Арутюнян, Е.Е. Дубинина, Н.Н. Зыбина - СПб. - 2000. - 67 с .
- Соколовский В.В. Тиолдисульфидное соотношение крови как показатель состояния неспецифической резистентности организма./ В.В. Соколовский. - СПб., 1996. - 30 с.
- Бресткин А.П., Фруентова Т.А. Влияние солей на холинэстеразный гидролиз ацетилхолина при различных значениях рН.// Биохимия.- 1968.- т. 3. - С. 817- 822.
- Окунь И.М., Лыскова Т.И., Аксенцева С.Л., Конев С.Н. //Влияние свободных жирных кислот на специфическое связывание уабаина Na,K-АТФ-азой в синаптических мембранах и синаптосомах мозга : Ионный гомеостаз и влияние факторов внешней среды на жизнедеятельность клетки. Из-во МГУ. -1987. - С. 86-87.
- Макаров В.Г. Онтогенетические и суточные особенности состояния тиолдисульфидной системы крови белых крыс // Флуктуации состояния биохимических систем. - Л., ЛСГМИ, 1986.- С. 12-18.
- Бойко Е.Р., Щадрина В.Д., Козловская А.В. Сезонные аспекты функционирования ферментативных антиоксидантных систем рожениц европейского севера. // Российский физиологический журнал им. И.М. Сеченова.- 2006.- т. 92, № 5. - С. 633-642.
- Шноль С.Э. Спонтанные обратимые изменения «конформационные колебания» препаратов мышечных белков. Автореф. докт. дис. Пущино-на-Оке, Ин-т биофизики АН СССР. 1970.
- Кулеба В.А., Тимофеева В.М. Избирательная чувствительность АТФ-аз к изменениям окислительно-восстановительного состояния среды // Антиоксиданты и адаптация. – Л., ЛСГМИ, 1979 .- С. 18-21.
- John D.Hayes, Timothy J. Mantle. Inhibition of hepatic and extrahepatic glutatyione - S-transferases by primary and secondary bile acids. // Biochem. J. -1986 v.233-P. 407-415.
Поиск



