
Возможность использования поведенческих ответов Danio rerio в оценке дозозависимых эффектов феназепама
Лебедев В.А., Лебедев А.А., Бычков Е.Р., Шабанов П.Д. Возможность использования поведенческих ответов Danio rerio в оценке дозозависимых эффектов феназепама. Лабораторные животные для научных исследований. 2018; 1. https://doi.org/10.29296/2618723X-2018-01-02
Резюме
Рассматривается возможность использования Danio rerio для оценки дозозависимых эффектов феназепама. Ранее у Danio rerio изучали действие анксиолитиков диазепама и хлордиазепоксида, но не феназепама. Использовали метод тест-стресса новизны: рыбу помещали сначала в мерный стакан с растворенным фармакологическим веществом (или водой), а затем – в просмотровый аквариум на 6 мин, где автоматически регистрировали траекторию движения, длину пути, число перемещений в верхнюю часть аквариума, время пребывания в нижней части аквариума, число и время паттерна «фризинг» за каждую минуту опыта. Результаты. Показано, что в ответ на новизну помещения в просмотровый аквариум рыбы реагируют погружением на дно, увеличением фризинга и снижением числа перемещений в верхнюю половину аквариума. На фоне введения феназепама рыбы находились уже не только в нижней, но и в верхней части просмотрового аквариума. Средняя длина пути существенно не изменялась в диапазоне использованных доз (0,1–1 мг/л). Число и время паттерна «фризинг», а также время пребывания в нижней части аквариума в эксперименте снижались более чем в 2 раза по сравнению с контролем и проявляли дозозависимый эффект. Число перемещений в верхнюю часть аквариума за опыт существенно увеличивалось: с 1 – в контроле до 57 – при использовании феназепама в дозе 1 мг/л. При анализе динамики параметров за каждую минуту показано, что время пребывания рыбы в нижней части аквариума снижалось, начиная с 3-й минуты опыта при использовании феназепама в дозе 0,5 мг/л. При этом число перемещений рыбы в верхнюю часть аквариума достоверно увеличивалось более чем в 2 раза, начиная с 3-й минуты опыта при использовании феназепама в дозе 1 мг/л. Заключение. Установлена высокая чувствительность тест-стресса новизны для изучения тревожно-фобических реакций у Danio rerio. Доказана перспективность использования Danio rerio в качестве животных моделей в поведенческой фармакологии, причем этот метод не уступает (а даже по ряду позиций превосходит) исследованиям на грызунах.
Введение
Исследования поведения Danio rerio (zebrafish) применяются главным образом в нейробиологии [15, 22], нейроэтологии [4, 16, 17] и нейрофармакологии [6, 14, 19]. Активное плавание, адаптация к новой обстановке, короткий репродуктивный период, высокая плодовитость и низкая себестоимость указывают на возможность использовать Danio rerio в качестве животных моделей для лабораторных исследований [25]. Тест стресса новизны используется для изучения тревожно-фобических реакций у Danio rerio. При этом животные демонстрируют стабильные поведенческие ответы под действием стресса новизны [3, 6, 17]. Данный поведенческий тест основывается на инстинкте поиска защиты от незнакомой обстановки погружением на дно, фризингом (обездвиживанием, или «примерзанием») и снижением двигательного (и исследовательского) поведения. После адаптации (акклиматизации) к новой обстановке двигательная активность увеличивается, уменьшается фризинг и увеличивается число переходов в верхнюю половину аквариума [20, 22, 28]. Подобное поведение традиционно анализируется в тесте «открытое поле» в исследованиях тревожности у грызунов [1, 2, 8, 10, 27]. А.В. Калуев и соавт. [9] применили показатели тревожности и адаптировали их непосредственно для Danio rerio. Рыбка помещается сначала в мерный стакан с растворенным фармакологическим веществом (или водой), а затем в небольшой просмотровый аквариум, где региструются двигательная активность, фризинг, число переходов в верхнюю половину аквариума и время в последней. В частности, было исследовано поведение Danio rerio после применения анксиолитиков и анксиогенов [26, 28], что коррелировало с уровнем кортизола в крови [12]. Поведенческие эффекты у Danio rerio показаны также после применения этанола [13], никотина [17], морфина [7], фенамина [23]), бензодиазепинов [3] и кокаина [19]. Влияние стресса новизны у Danio rerio можно тестировать и в ряде дополнительных тестов, таких как свет-темнота [21, 23], Т-образный лабиринт [24], открытое поле [18]. Таким образом, можно более подробно проанализировать поведение животных. Кроме того, существует ряд тестов и для неполовозрелых животных [4, 11]. В последние годы для регистрации поведения Danio rerio применяют компьютерный анализ видеотрека, что также позволяет более детально, с высокой точностью изучить эффекты фармакологических веществ [6, 12, 13].
Цель настоящей работы – оценка возможности использования Danio rerio для анализа дозозависимых эффектов феназепама. В исследованиях уровня тревожности и влияния стресса новизны на поведенческие ответы у Danio rerio ранее показано действие диазепама и хлордиазепоксида [3, 10], наиболее известных транк-вилизаторов, которые применяются в клинике многих стран. При этом отечественный препарат феназепам, обладающий более мощным анксиолитическим действием, ранее не был исследован у Danio rerio. Поэтому настоящее исследование было посвящено анализу действия анксиолитика феназепама у Danio rerio.
Методы исследования
Выбор животных. В работе использованы 104 молодые половозрелые рыбы Danio rerio (zebrafish) в возрасте 6–8 мес (жизненный цикл до 5 лет) фирмы «Аква Питер», выращенные в Институте экспериментальной медицины (ИЭМ). Danio rerio – дикий тип (wild type). Для тестирования использовали интактных животных после 2-недельного периода адаптации к помещению в аквариум водоизмещением 40 л по 20–30 рыб в каждом. Температуру воды 25–27°С поддерживали постоянно. Животных содержали в стандартных условиях светового режима 8.00–20.00 ч при температуре помещения 22±2°С, кормили 2 раза в день стандартным кормом tetramin tropical flakes.
Тест стресса новизны. Для экспериментов оценки новизны применяли стандартный просмотровый аквариум, который используется для изучения тревожно-фобических реакций у Danio rerio [6]. Он имел водоизмещение 1,5 л, трапециевидную форму, высоту 15 см и ширину 7 см. Длина аквариума в основании составляла 22 см, а в верхней части – 28 см. Такая конструкция аквариума позволяет минимизировать латеральные движения рыбки и свободно наблюдать за вертикальными и горизонтальными движениями. Поскольку данный поведенческий тест тест основывается главным образом на инстинкте поиска защиты от незнакомой обстановки погружением на дно [12, 16], аквариум был разделен чертой на 2 равные части – верхнюю и нижнюю. Рыбка помещалась сначала в мерный стакан водоизмещением 200 мл с растворенным фармакологическим веществом (или водой) на 5 мин, затем – в предстартовый аквариум с водой (10×10×10 см3) на 5 мин и далее – в просмотровый аквариум на 6 мин, где регистровали двигательную активность во время опыта (длина трека рыбки), число переходов в верхнюю и нижнюю половины аквариума и время нахождения в них последовательно за каждую минуту опыта. Автоматически регистрировали число и время паттернов «фризинг» (обездвиживание, или «примерзание») за опыт, которые обычно наблюдаются при стрессе новизны и отражают уровень тревожности животного. Поведение регистрировали автоматически с помощью системы Noldus EthoVision XT7, которая позволяет просмотреть видеотреки рыбки. Система снимает показания как в цифровом выражении, так и визуально, а также контролирует видеотрек. Весь период наблюдения непрерывно регистрировался, при помощи программы шла запись траектории движения (см. рисунок).

Фармакологические вещества. Для фармакологического анализа использовали субстанцию феназепама ОАО «Новосибхимфарм» (Россия). Феназепам растворяли в воде для аквариумов и применяли в 3 дозах: 1) 0,1 мг на 1000 мл воды; 2) 0,5 мг на 1000 мл воды 3) 1 мг на 1000 мл воды.
Статистическая обработка полученных данные. Применяли пакеты Stаstistica v.6
и Origin v.7.5. Нормальность распределений оценивали при помощи критерия Колмогорова–Смирнова, значимость
различий между группами – при помощи
t-критерия Стьюдента либо U-критерия Манна–Уитни. Данные
представлены в виде «среднее арифметическое ± среднеквадратичное отклонение».
Результаты исследований
В процессе проведения экспериментов были установлены типичные поведенческие реакции на новизну (т.е. на помещение животного в просмотровый аквариум). Как правило, рыба находилась в это время в нижней части просмотрового аквариума и совершала небольшие перемещения вдоль него (см. рисунок). Средняя длина пути рыбы, который автоматически регистрировался по продолжительности трека, составляла 1205,0±91,8 см (табл. 1). При этом часто наблюдали фризинг («примерзание», время полной неподвижности). Число и время паттерна «фризинг» автоматически регистрировались и составляли за опыт соответственно 68,2±5,7 и 36,5±4,5 с. Среднее время пребывания рыбы в нижней части аквариума составляло 351,0±36,4 с. Динамика времени пребывания рыбы в нижней части аквариума регистрировалась автоматически и существенно не изменялась (табл. 2). Число перемещений рыбы в верхнюю часть просмотрового аквариума за опыт составляло 1,2±0,8. Динамика числа перемещений рыбы в верхнюю часть аквариума также регистрировалась автоматически и существенно не изменялась (табл. 3).
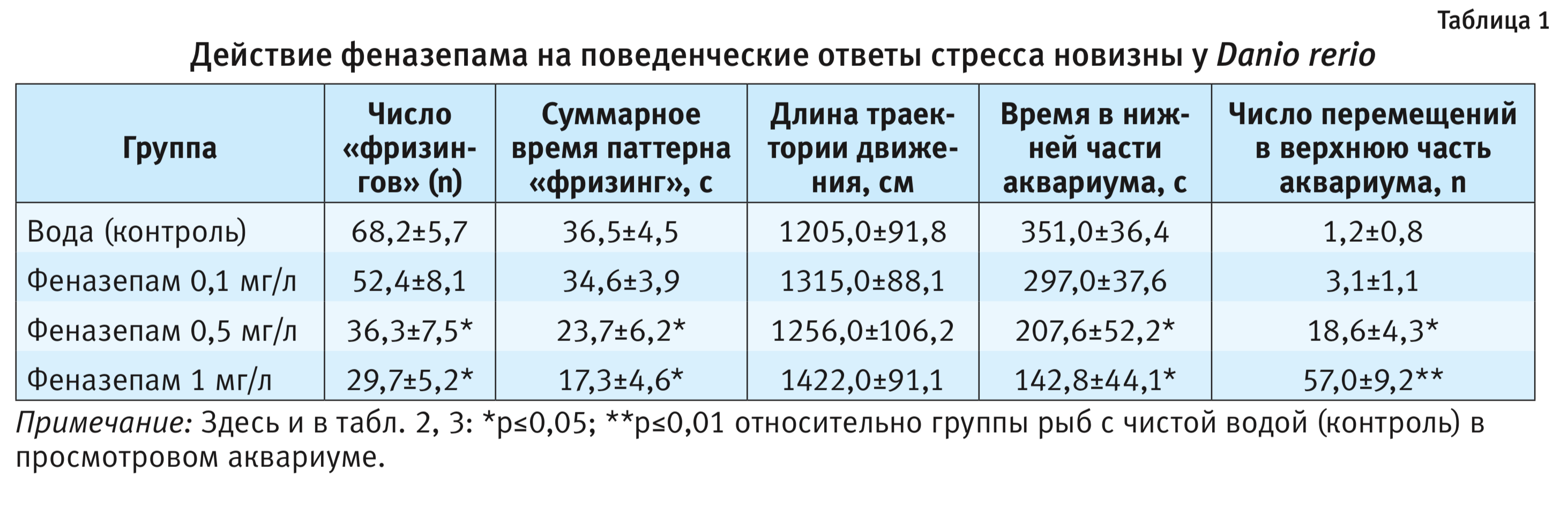
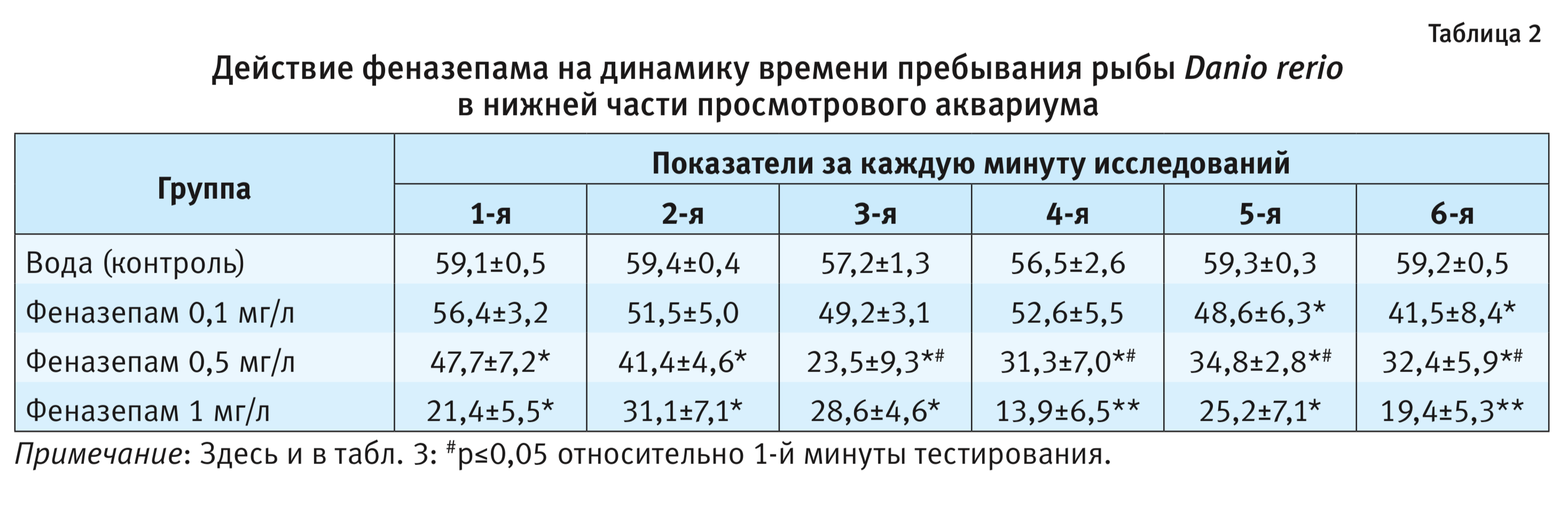
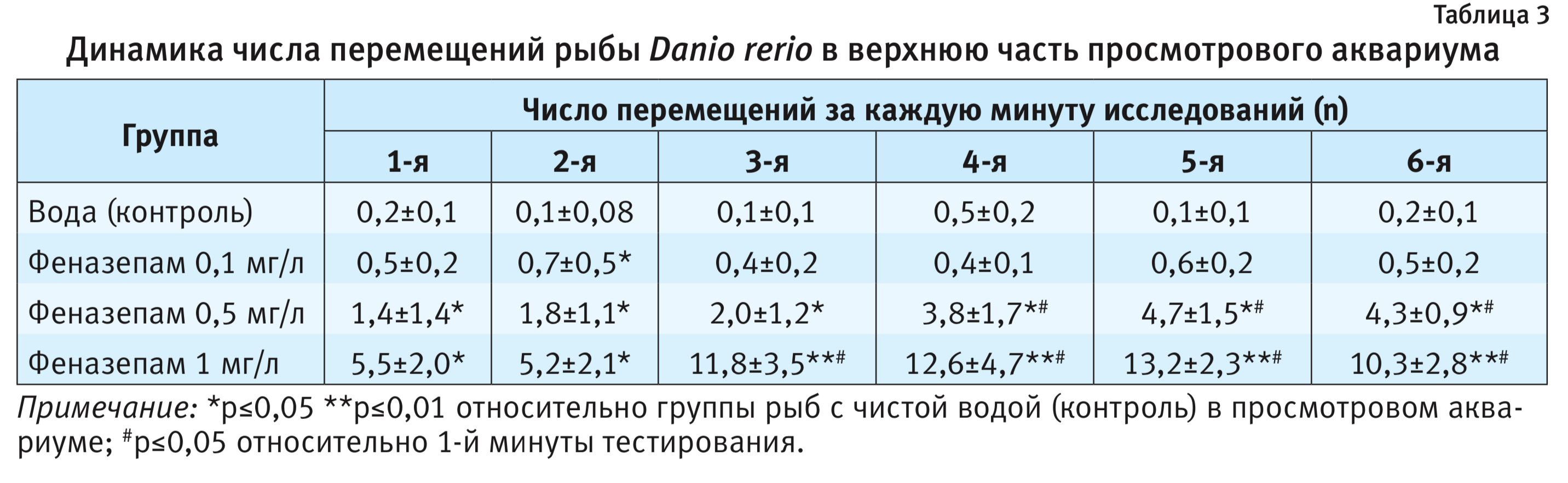
На фоне введения феназепама были выявлены типичные для анксиолитика поведенческие реакции на новизну при помещении животного в просмотровый аквариум. Как правило, рыбы находились в это время не только в нижней части просмотрового аквариума, но часто заплывали и в верхнюю его часть (см. рисунок). На фоне феназепама средняя длина пути рыбы существенно не изменялась в диапазоне использованных доз по сравнению с контролем. При этом так же, как и в контроле, наблюдался фризинг. Число и время паттерна «фризинг» достоверно снижались по сравнению с контрольной группой животных (р<0,05) и проявляли дозозависимый эффект. Среднее время пребывания рыбы в нижней части аквариума также достоверно снижалось по сравнению с контрольной группой животных (р<0,05) и составляло при введении 0,1; 0,5 и 1 мг/л соответственно 297,0±37,6; 207,6±52,2 и 142,8±44,1, проявляя неизменно дозозависимый эффект (см. табл. 1). Время пребывания рыбы в нижнем отсеке снижалось, начиная с 3-й минуты опыта при использовании феназепама в дозе 0,5 мг/л по сравнению с 1-й минуты тестирования (р<0,05). Число перемещений рыбы в верхнюю часть просмотрового аквариума за опыт достоверно увеличивалось по сравнению с контролем (р<0,01) и составляло при введении 0,1; 0,5 и 1 мг/л соответственно 3,1±1,1, 18,6±4,3 и 57,0±11,2. Число перемещений рыбы в верхнюю часть просмотрового аквариума достоверно увеличивалось, начиная с 3-й минуты опыта при использовании феназепама в дозе 1 мг/л (см. табл. 3). Время пребывания рыбы в нижней части просмотрового аквариума достоверно увеличивалось начиная с 3-й минуты опыта при использовании феназепама в дозе 0,5 мг/л по сравнению с 1-й минутой тестирования.
Обсуждение полученных результатов. Настоящие исследования показали, что реакция на новизну помещения в просмотровый аквариум у Danio rerio (zebrafish) имеет типичную картину или типичные паттерны поведения. В ответ на незнакомую обстановку просмотрового аквариума рыба реагировала погружением на дно, фризингом (обездвиживанием или «примерзанием») и снижением двигательного поведения. Как правило, рыба большую часть времени находилась в нижней части просмотрового аквариума и совершала небольшие перемещения вдоль него (см. рисунок). При этом часто наблюдали фризинг, число и время которого за опыт было достаточно велико, как и время пребывания рыбы в нижней части аквариума. Динамика времени пребывания рыбы в нижнем отсеке практически не изменялась, так же, как и таковая пребывания рыбы в верхнем отсеке и динамика числа перемещений рыбы в верхний отсек.
Полученные результаты во многом соответствуют данным литературы, где отмечается, что тестирование стресса новизны применяется в ряде лабораторий для исследования уровня тревожности у Danio rerio, и рыбы показывают, как правило, типичную картину поведенческих паттернов [3, 6, 17]. При этом рыбы плавают в нижней части аквариума и часто замирают (демонстрируя фризинг). Двигательное (и исследовательское) поведение значительно снижается по сравнению с поведением в домашнем аквариуме [26, 28]. После периода адаптации к просмотровому аквариуму поведение становится во многом подобно таковому в домашнем аквариуме, т.е. двигательная активность увеличивается, уменьшается фризинг и увеличивается число переходов в верхнюю половину аквариума [17, 18, 24].
Кроме того, реакция на новизну помещения в просмотровый аквариум у Danio rerio имеет типичную картину паттернов поведения, которая меняется на фоне введения феназепама. Метод регистрации поведения оказался высокочувствительным к анксиолитическому действию исследуемого вещества. При этом рыбы плавали в это время не только в нижней части просмотрового аквариума, но и стали активнее подниматься вверх, а к концу опыта проводили здесь значительную часть времени (см. рисунок). Длина траектории рыбы после введения феназепама существенно не увеличивалась по сравнению с контрольной группой животных (см. табл. 1). Число и время паттерна «фризинг» после введения феназепама достоверно снижались по сравнению с контрольной группой животных, при этом регистрировали дозозависимый эффект, что весьма важно. Время пребывания рыбы в нижней части аквариума после введения феназепама значительно снижалось, особенно при использовании в дозе 0,5 и 1 мг/л. Показатели динамики времени пребывания рыбы в нижнем отсеке также снижались, начиная с 3-й минуты опыта по сравнению с контролем, особенно при использовании феназепама в дозе 0,5 мг/л. Число перемещений рыбы в верхнюю часть просмотрового аквариума после введения феназепама достоверно увеличивалось по сравнению с контролем, особенно при использовании препарата в дозе 1 мг/л. При этом динамика числа перемещений рыбы в верхнюю часть просмотрового аквариума также менялась, показатели достоверно увеличивались, начиная с 3-й минуты опыта, особенно при использовании препарата в дозе 0,5 и 1 мг/л.
Результаты исследования не противоречат данным литературы. Так, тестирование анксиолитиков в ряде лабораторий с целью изучения противотревожного эффекта у Danio rerio показало, что рыбы демонстрируют достаточно типичную картину поведенческих паттернов, аналогичную полученным результатам [3, 6, 17]. При этом паттерны поведения под действием транквилизаторов у Danio rerio сходны с таковыми у грызунов (крысы, мыши) – наиболее частом объекте изучения транквилизаторов.
В настоящей работе был проведен фармакологический анализ действия бензодиазепинового транквилизатора феназепама, что ранее не исследовалось. В литературе описаны лишь анксиолитические эффекты диазепама [10] и хлордиазепоксида [3], наиболее известных транквилизаторов, широко применяемых во многих странах. Следует сказать, что в указанных работах не отмечено принципиальных различий в эффектах этих транквилизаторов, более того, сделан вывод о том, что бензодиазепиновые анксиолитики диазепам и хлордиазепоксид не уступают по выраженности противотревожного эффекта агонисту 5HT1A рецепторов буспирону. Единственным отличием в действии бензодиазепиновых препаратов авторы называют пороговую анксиолитическую дозу, на которую Danio rerio реагируют, что не вызывает седативного эффекта, т.е. существенного снижения двигательной активности [3]. В наших опытах выявлена не только основная квалификационная характеристика отечественного транквилизатора феназепама, заключающаяся в снятии стресса новизны без изменения двигательной активности (типичный анксиолитический эффект, зависимый от дозы), но и дополнительные характеристики, к которым относится изменение характера двигательной активности между верхней и нижней частями просмотрового аквариума за счет возрастания его до гиперактивности в верхней части аквариума, где в норме рыбы, как правило, проводят чрезвычайно мало времени. При этом в нижней части аквариума двигательная активность снижалась умеренно, максимально – в 2 раза, после введения феназепама в большой (1 мг/л) дозе, соответствующей 0,0001% раствору препарата. Эти характеристики, безусловно могут быть использованы и при оценке других классов (групп) лекарственных препаратов, что открывает определенные перспективы использования Danio rerio в качестве недорогого, но достаточно информативного теста для целей фармакологии.
Заключение
На основании полученных данных можно сделать выводы:
- Тест-стресса новизны является весьма чувствительным для изучения тревожно-фобических реакций у Danio rerio. Использования Danio rerio в качестве животной модели в поведенческой фармакологии перспективно и не уступает (а даже по ряду позиций превосходит) исследованиям на грызунах.
- Фармакологический анализ анксиолитического действия феназепама у Danio rerio показал, что феназепам дозозависимо снижает (по сравнению с контрольной группой рыб) число и время фризинга, увеличивает число перемещений в верхнюю половину аквариума и время плавания в последней.
Список источников
- Лебедев А.А., Пшеничная А.Г., Бычков Е.Р., Якушина Н.Д., Шабанов П.Д. Антагонист рецепторов кортиколиберина астрессин снимает тревожно-фобические состояния у крыс, выращенных в социальной изоляции. Обзор по клин. фармакол. и лек. терапии. 2016. Т. 14. № 4: 24–31 [Lebedev A.A., Pshenichnaya A.G., Bychkov E.R., Yakushina N.D., Shabanov P.D. Antagonist receptorov kortikoliberina astressin snimaet trevozhno-fobicheskie sostoyaniya u krys, vyrashchennyh v social’noj izolyacii. Obzory po klinicheskoj farmakologii i lekarstvennoj terapii. 2016; 14 (4): 24–31 (in Rus)].
- Шабанов П.Д., Лебедев А.А., Якушина Н.Д., Пшеничная А.Г., Бычков Е.Р. Моделирование обсессивно-компульсивного и аддиктивного игрового поведения у крыс введением фенамина в тесте закапывания шариков. Наркология. 2017. Т. 16; 1 (181): 32–8 [Shabanov P.D., Lebedev A.A., Yakushina N.D., Pshenichnaya A.G., Bychkov E.R. Modelirovanie obsessivno-kompul'sivnogo i addiktivnogo igrovogo povedeniya u krys vvedeniem fenamina v teste zakapyvaniya sharikov. Narkologiya. 2017; 1 (181): 32–8 (in Rus)].
- Bencan Z., Sledge D., Levin E.D. Buspirone, chlordiazepoxide and diazepam effects in a Danio rerio model of anxiety. Pharmacol. Biochem. Behav. 2009; 94 : 75–80. DOI: 10.1016/j.pbb.2009.07.009.
- Best J.D., Alderton W.K. Danio rerio: an in vivo model for the study of neurological diseases. Neuropsychiatr Dis Treat. 2008; 4: 567–76.
- Best J.D. Non-associative learning in larval Danio rerio. Neuropsychopharmacology. 2008; 33: 1206–15. DOI: 10.1038/sj.npp.1301489
- Blaser R., Gerlai R. Behavioral phenotyping in Danio rerio: comparison of three behavioral quantification methods. Behav Res Methods. 2006; 38: 456–69.
- Bretaud S. A choice behavior for morphine reveals experience-dependent drug preference and underlying neural substrates in developing larval Danio rerio. Neuroscience. 2007; 146: 1109–16. DOI: 10.1016/j.neuroscience.2006.12.073
- Bronikowski A.M. Open-field behavior of house mice selectively bred for high voluntary wheel-running. Behav Genet. 2001; 31: 309–16.
- Cachat J., Stewart A., Grossman L., Kalueff A.V. Measuring behavioral and endocrine responses to novelty stress in adult Danio rerio. Nature protocols. 2010; 5 (11): 1786–9. DOI: 10.1038/nprot.
- Choleris E, Thomas AW, Kavaliers M, Prato FS. A detailed ethological analysis of the mouse open field test: effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neurosci Biobehav Rev. 2001; 25: 235–60.
- Creton R. Automated analysis of behavior in Danio rerio larvae. Behav Brain Res. 2009; 203: 127–36. DOI:10.1016/j.bbr.2009.04.030
- Egan R.J. Understanding behavioral and physiological phenotypes of stress and anxiety in Danio rerio. Behav Brain Res. 2009; 205: 38–44. DOI: 10.1016/j.bbr.
- Gerlai R. Event recording and video-tracking: towards the development of high-throughput Danio rerio screens. In Proc. 5th Conf. on Methods in Behav. Res. Eds. J.J. Noldus, F. Grieco, L.W.S. Loijens, P.H. Zimmerman. 2005.
- Gerlai R., Lee V., Blaser R. Effects of acute and chronic ethanol exposure on the behavior of adult Danio rerio (Danio rerio). Pharmacol Biochem Behav. 2006; 85: 752-761. DOI: 10.1016/j.pbb.2006.11.010
- Guo S. Using Danio rerio to assess the impact of drugs on neural development and function. Expert Opin Drug Discov. 2009; 4: 715–26. DOI: 10.1517/17460440902988464
- Key B., Devin C.A. Danio rerio as an experimental model: strategies for developmental and molecular neurobiology studies. Methods Cell Sci. 2003; 25: 1–6.
- Levin E.D., Chen E. Nicotinic involvement in memory function in Danio rerio. Neurotoxicol Teratol. 2004; 26: 731–5. DOI:10.1016/j.ntt.2004.06.010
- Levin E.D., Bencan Z., Cerutti D.T. Anxiolytic effects of nicotine in Danio rerio. Physiol Behav. 2007; 90: 54–8. DOI: 10.1016/j.physbeh.2006.08.026
- Lopez-Patino M.A., Cabral H., Zhdanova I.V. Anxiogenic effects of cocaine withdrawal in Danio rerio. Physiol Behav. 2008; 93: 160–71. DOI: 10.1016/j.physbeh.2007.08.013
- Maximino C., Marques B.T., Dias C.A., Gouveia A., Morato S. Parametric analyses of anxiety in Danio rerio scototaxis. Behav. Brain Res. 2010; 210: 1–7. DOI: 10.1016/j.bbr.2010.01.031
- Maximino C., Marques B. T., Dias C.A., Gouveia A., Morato S. Scototaxis as anxiety-like behavior in fish. Nat Protoc. 2010; 5: 209–16. DOI: 10.1038/nprot.2009.225
- Miklosi A., Andrew R.J. The Danio rerio as a model for behavioral studies Danio rerio. – 2006; 3: 227–34. DOI: 10.1089/zeb.2006.3.227
- Ninkovic J., Bally-Cuif L. The Danio rerio as a model system for assessing the reinforcing properties of drugs of abuse. Methods. 2006; 39: 262–74. DOI:10.1016/j.ymeth.2005.12.007
- Sackerman J. Danio rerio behavior in novel environments: effects of acute exposure to anxiolytic compounds and choice of Danio rerio line. Int J Comp Psychol. 2010; 23: 43–61.
- Spence R., Gerlach G., Lawrence C., Smith C. The behaviour and ecology of the Danio rerio, Danio rerio. Biol. Rev. Camb. Philos. Soc. 2008; 83: 13–34. DOI: 10.1111/j.1469-185X.2007.00030.x
- Stewart A. The developing utility of Danio rerio in modeling neurobehavioral disorders. Int J Comp Psychol. 2010; 23: 104–21. DOI: 10.1016/j.pnpbp.2010.11.035
- Streng J. Open-field behavior in four inbred mouse strains. Can. J. Psychol. 1971; 25: 62–8.
- Wong K. Analyzing habituation responses to novelty in Danio rerio (Danio rerio). Behav. Brain Res. 2009; 208: 457. DOI:10.1016/j.bbr.2009.12.023
Поиск



