Сравнительная морфология поджелудочной железы экспериментальных животных и человека
Гущин Я.И., Шедько В.В., Мужикян А.А., Макарова М.Н., Макаров В.Г. Сравнительная морфология поджелудочной железы экспериментальных животных и человека. Лабораторные животные для научных исследований. 2018; 3. https://doi.org/10.29296/2618723X-2018-03-04
Резюме
Поджелудочная железа – непарный паренхиматозный орган относится к железам смешанной секреции, включает в себя экзокринную и эндокринную части, участвует в пищеварении и регуляции углеводного, белкового и жирового обменов в тканях. Работа поджелудочной железы подчиняется тонкой нейро-гуморальной регуляции, ее патология приводит к серьезным последствиям. В доклинических исследованиях используются разные виды лабораторных животных, поджелудочная железа которых по своему строению и функции чрезвычайно схожа с человеческой. Однако есть и отличия, которые могут существенно влиять на результат эксперимента. Данное сравнительное исследование было направлено на определение схожести и различия в морфологии органа у человека и лабораторных животных. Согласно анализу данных литературы, выявлены основные черты сходства строения поджелудочной железы человека и лабораторных животных, как в анатомическом, так и гистологическом строении. Железа имеет одинаковые источник развития и закономерности органо- и гистогенеза. Выраженно отличается анатомическое строение железы: у человека – компактного типа, у рассматриваемых животных – скорее брыжеечного типа. Хотя мнения авторов по данному вопросу отличаются. Несмотря на разнообразие формы у каждого вида, в железе выделяют головку, тело и хвост. Количество протоков у разных животных также вызывает споры исследователей. В литературе можно найти описание нескольких вариантов протоковой системы у животных. Гистологическое строение железы также однотипно. Экзокринная часть не имеет существенных особенностей у разных видов. А в строении островкового аппарата можно выделить видовые различия в качественном и количественном составе инсулоцитов в островках Лангерганса. Данная статья может быть полезна не только фармакологам и токсикологам, работающим в области доклинических исследований, но и гистологам, физиологам и патологам, изучающих патологические процессы и механизмы функционирования поджелудочной железы.
Введение
Поджелудочная железа (pancreas) – непарный паренхиматозный орган, расположенный позади желудка, преимущественно ретроперитонеально (только передняя ее поверхность покрыта брюшиной). Она относится к железам смешанной секреции, поскольку включает в себя экзокринную и эндокринную части. В экзокринной части продуцируется панкреатический сок, богатый пищеварительными ферментами (трипсин, липаза, амилаза и пр.), а в эндокринной части вырабатываются различные гормоны (инсулин, глюкагон, соматостатин, (вазоактивный интестинальный полипептид – ВИП), панкреатический полипептид). Железа участвует в пищеварении, вырабатывая пищеварительные ферменты, поступающие в двенадцатиперстную кишку и осуществляющие гидролиз практически всех составных частей пищи, поступающей в организм [1]. Деятельность органа чрезвычайно важна для регуляции углеводного (энергетического), белкового и жирового обменов в тканях животных и человека [2–6]. Работа поджелудочной железы подчиняется тонкой нейрогуморальной регуляции, ее патология приводит к серьезным последствиям.
В доклинических исследованиях используются разные виды лабораторных животных, поджелудочная железа которых по своему строению и функции чрезвычайно схожа с человеческой. Однако есть и отличия, которые могут существенно влиять на результат эксперимента. Задача данного сравнительного исследования – выявить схожесть и различия в морфологии органа у человека и лабораторных животных. Такие знания могут быть полезны не только фармакологам и токсикологам, работающим в области доклинических исследований, но и для гистологов, физиологов и патологов, изучающих патологические процессы и механизмы функционирования поджелудочной железы.
Развитие поджелудочной железы
О развитии поджелудочной железы, как человека, так и лабораторных животных (преимущественно грызунов и, особенно, крыс), написано много работ, однако в них встречается число противоречий.
Установлено, что по источнику развития и закономерностям органо- и гистогенеза поджелудочная железа человека и белой крысы схожи [7, 8].
Закладка железы начинается у человека в конце 3-й начале 4-й недели эмбрионального развития, а у крыс – на 10–11-е сутки. Она из энтодермы и мезодермы и имеет несколько зачатков. У человека выделяют 2 зачатка – вентральный и дорзальный [2, 8], в то время как у животных выделяют 3 зачатка – 2 вентральных и дорзальный [7]. Однако есть авторы, которые выделяют у крыс только 2 зачатка [8].
Вентральный зачаток поджелудочной железы представляет собой выпячивание скопления энтодермы зародышевой средней кишки в толщу мезенхимы между листками вентральной брыжейки. Дорсальный зачаток поджелудочной железы, зарождающийся раньше, располагается каудальнее вентрального зачатка, является выпячиванием печеночного дивертикула и первичной кишечной трубки в толщу мезенхимы дорсальной брыжейки. Оба зачатка сближаются и сливаются (у человека на 6–7-й неделе [7] и на 2-й неделе – у крыс [8]), формируя единую закладку, которая располагается в толще мезенхимы дорсальной брыжейки. Затем происходит канализация (тубуляция) эпителиальных тяжей: они приобретают форму трубочек, стенки которых представлены одним слоем цилиндрического эпителия; их количество постепенно нарастает. На их боковых стенках и концах появляются выпячивания в виде почек – зачатки концевых отделов – ацинусов, стенка которых образована одним слоем цилиндрического (призматического) эпителия. Плотно прилежащие друг к другу концевые отделы, сгруппированные в скопления, формируют первичные дольки, отграниченные тонкими прослойками рыхлой соединительной ткани.
Примерно на 3-м месяце внутриутробного развития у человека [2, 8] и на 15–16-е сутки – у крыс [8] происходит формирование экзокринной части железы за счет дифференцировки островковых клеток предшественниц, клеток эпителия протоков, преобразования ацинарных клеток и пролиферации дифференцированных эндокринных клеток в островках [9]. Образованные островки располагаются в толще долек и, как правило, тесно контактируют с концевыми отделами или выводными протоками. Во 2-й половине пренатального периода поджелудочная железа имеет отчетливое дольчатое строение. После рождения гистогенез поджелудочной железы не завершается – по периферии органа можно обнаружить эпителиальные трубочки с выпячиваниями, в которых продолжается активное деление клеток и формирование ткани железы [10], что связано с изменением условий функционирования организма.
Анатомия
Форма поджелудочной железы у человека и животных крайне разнообразна. Выделяют несколько типов строения: компактный, рассеянный или брыжеечный и промежуточный, сочетающий оба варианта [11]. У человека железа представляет собой компактный орган [2, 11, 12]. Большинство авторов сходятся во мнении, что поджелудочная железа крысы, мыши, кролика, морской свинки и хомяка классифицируется по брыжеечному типу. При таком типе ткани поджелудочной железы диффузно распределены в брыжейке двенадцатиперстной кишки, поперечной ободочной кишки и большого сальника, прилегающего к желудку и селезенке [3, 5, 13–17]. Так, у крысы, а в еще большей степени – у морской свинки, поджелудочная железа отличается своей рыхлостью, изогнутостью и разветвленностью [5, 15, 18, 19], так как состоит из отдельных маленьких гроздевидных долек, связанных рыхлой соединительной тканью в одно древовидно разветвляющееся образование [3, 14].
Однако Minoru Tsuchitani (2016) классифицирует поджелудочную железу крысы и мыши как промежуточный тип, поскольку селезеночная часть у указанных животных относительно компактна, но часть диспергирована в брыжейке. Он относит поджелудочную железу хомяков к 1-му – концентрированному типу. Поджелудочная железа кролика относительно мала. Она располагается в брыжейке между поперечной ободочной кишкой, желудком и двенадцатиперстной кишкой [20, 21] и состоит из отдельных частей неправильной формы, которые размещены в перитонеальной складке [22]. Дольки имеют, как правило, яйцевидную форму, 2–3 мм в диаметре и около 5 мм толщиной [23]. По мнению большинства авторов, такое строение – рассеянный, брыжеечный тип [11, 24]. Однако В.Н. Жеденов (1957) допускает существование как рассеянного, так и концентрированного типа, при котором ткань собрана компактно и напоминает единый орган [25]. У морских свинок форма поджелудочной железы в брюшной полости имеет S-образную форму, схожую с таковой у человека, но в расправленном состоянии напоминает бабочку [15]. Головка поджелудочной железы почти полностью окружена двенадцатиперстной кишкой и отграничена от желудка луковицей двенадцатиперстной кишки [15].
Поджелудочная железа человека характеризуется компактным расположением и лежит в подкове двенадцатиперстной кишки, на уровне I–II поясничных позвонков и покрыта тонкой соединительнотканной капсулой. Позади нее находится позвоночник, аорта, нижняя полая и левая почечная вены, спереди располагается желудок [12]. У взрослого человека длина ее составляет 14–20 см, ширина 3–9 см, толщина 2–3 см, масса 70–100 г [34, 39]. В поджелудочной железе выделяют головку, тело и хвост.
Головка (caput pancreatis) охвачена двенадцатиперстной кишкой, задняя поверхность прилежит к правой почечной и воротной венам, передняя – к поперечной ободочной кишке. На границе с телом имеется вырезка (incisura pancreatis), в которой проходят верхняя брыжеечная артерия и вена.
Тело (corpus pancreatis) трапециевидной формы, и имеет заднюю поверхность и покрытые брюшиной нижнюю и переднюю поверхности. На передней поверхности выделяется выпуклость в сторону сальника – сальниковый бугор (tuber omentale), который обнаруживается только у человека и хомяка, но в области хвоста есть железы [26].
Хвост (cauda pancreatis) направляется влево и лежит рядом с селезенкой, левым надпочечником и левой почкой.
Несмотря на разнообразное строение, в поджелудочной железе лабораторных животных, как и в поджелудочной железе человека, условно выделяют 3 основные части – головка (дуоденальная часть), тело (пилорическая часть) и хвост (желудочно-селезеночная часть) [3, 16]. По данным Minoru Tsuchitani (2016), поджелудочная железа крысы разделена на 4 доли: желудочный, селезеночный, парабилиарный и двенадцатиперстный сегменты (именуемые также лопастями [11, 16, 27]), которые формируют 2 крайние формы строения и топографии – молоткообразную и трилистника, когда доли вдаются в смежные брыжейки.
Таким образом, расположение частей железы следующее: парабилиарный сегмент и двенадцатиперстный сегмент расположена на дуоденальной стороне, желудочно-селезеночный сегмент простирается до желудка и селезенки, концевая часть селезеночного сегмента заканчивается около ворот селезенки, каудальная часть двенадцатиперстного сегмента и дорсальная часть селезеночного сегмента часто соединены вместе и простираются до ободочной кишки (рис. 1) [11].
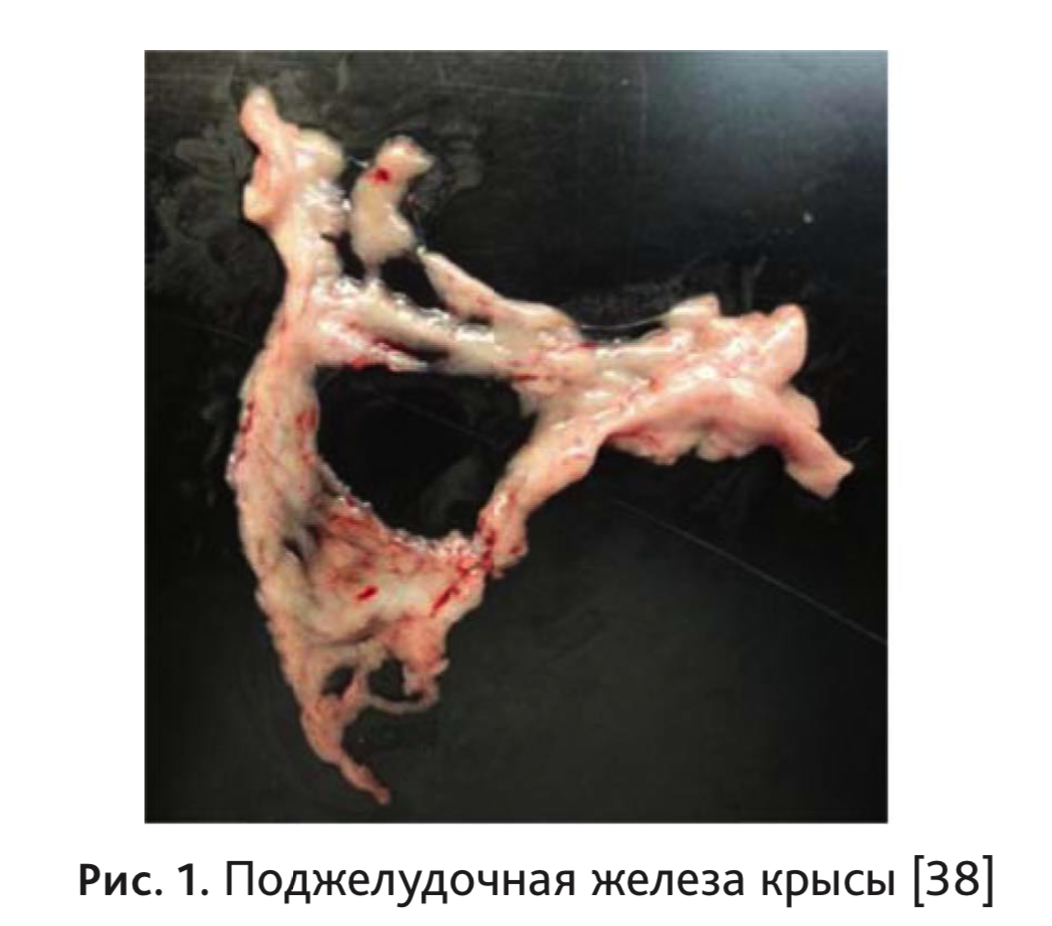
Протоки поджелудочной железы. У человека выделяют главный проток поджелудочной железы (ductus pancreaticus) или Вирсунгов проток, который принимает многочисленные ветви и, соединившись с общим желчным протоком, открывается общим большим сосочком двенадцатиперстной кишки (papilla duodeni major). Кроме того существует анатомически вариабельный добавочный или Санториниев проток (ductus pancreaticus accesorius), который начинается от головки поджелудочной железы и открывается в полость двенадцатиперстной кишки малым дуоденальным сосочком (papilla duodeni minor) [28].
На сегодняшний день мнения ученых о количестве выводных протоков поджелудочной железы у лабораторных
животных расходятся. Так, по данным
М.Р. Бектемировой, особенностью строения поджелудочной железы крысы
является наличие 2, очень тонких, выводных протоков, которые впадают либо в желчный проток, либо
непосредственно в двенадцатиперстную кишку [3]. В литературе встречаются данные о наличии у крыс
нескольких протоков поджелудочной железы, которые на различных уровнях впадают в общий желчный проток. По
мнению ряда исследователей, у крыс, как и у мышей, общий желчный проток открывается
в двенадцатиперстной кишке в большой сосочек двенадцатиперстной кишки. При этом отмечается, что
у крыс, а также у морских свинок отсутствует малый сосочек [29, 30].
Кроме того, у мышей также есть 1 или несколько небольших дополнительных протоков поджелудочной железы, которые высвобождают содержимое непосредственно в двенадцатиперстную кишку, оканчиваясь при этом одним или несколькими малыми сосочками двенадцатиперстной кишки соответственно. K. Vashisht (2015) делает вывод, что анатомия выводных протоков поджелудочной железы у крыс и мышей несколько отличается.
Согласно другим исследованиям, у крыс и мышей междольковые протоки открываются в общий печеночный проток, который входит в двенадцатиперстную кишку, а также есть некоторые протоки, которые открываются непосредственно в двенадцатиперстную кишку [17], имеется и добавочный проток, который всегда впадает в кишку самостоятельно [31].
У кроликов главный панкреатический проток открывается около конца двенадцатиперстной кишки, вдали от входа желчных протоков, на расстоянии в 30–50 см от привратника [20, 21, 23]. Кроме того, есть добавочный проток поджелудочной железы, открывающийся самостоятельно в двенадцатиперстную кишку [20, 21]. Ученые сходятся во мнении, что добавочные протоки могут полноценно заменить собой главный в случае закупорки или экспериментальной перевязки главного [20, 21, 30]. Хотя по данным C.A. Johnson-Delaney (2006), существует 1 панкреатический проток, который дренирует обе доли и открывается на стыке поперечной и восходящей петли двенадцатиперстной кишки. При этом данный проток при эмбриональном развитии был добавочным, но впоследствии остался единственным, поскольку произошла облитерация терминальной части главного [32].
Кровоснабжение железы у человека и животных очень схожи [12, 25, 33–35]. Поджелудочная железа питается из бассейна чревного ствола ветвями печеночной артерии – желудочно-двенадцатиперстными артериями (arteriae pancreaticoduodenales), а также ветвями селезеночной артерии (arteria lienalis). Венозный отток происходит по одноименным венам в систему воротной вены.
Иннервация поджелудочной железы также однотипна. Она имеет симпатическую, парасимпатическую и автономную иннервацию. Парасимпатическая часть нервной системы регулирует секрецию гормонов посредством ветвей блуждающего нерва (nervus vagus) [12, 36]. Симпатические тела нервных клеток расположены в брыжеечных и брюшнополостных ганглиях.
Автономная нервная система, как у человека, так и у животных, распространена по всей железе в соединительнотканных перегородках и паренхиме [37, 38]. Интересно ее взаиморасположение с островковым аппаратом, поскольку вместе с инсулоцитами образуются нейроинсулярные комплексы 2 типов: I тип – смесь островковых и ганглиозных клеток, при типе II нервные волокна лежат рядом с инсулярными клетками [39].
Гистология
Снаружи железа покрыта тонкой соединительнотканной капсулой. Паренхима органа разделена на дольки (рис. 2), между которыми располагаются перегородки или септы – соединительнотканные тяжи с проходящими в них кровеносными сосудами, ганглиями и нервами, а также выводными протоками. Поджелудочная железа мыши, в отличие от других грызунов, имеет толстую капсулу из плотной волокнистой неоформленной соединительной ткани, сросшейся с висцеральным листком брюшины [40].
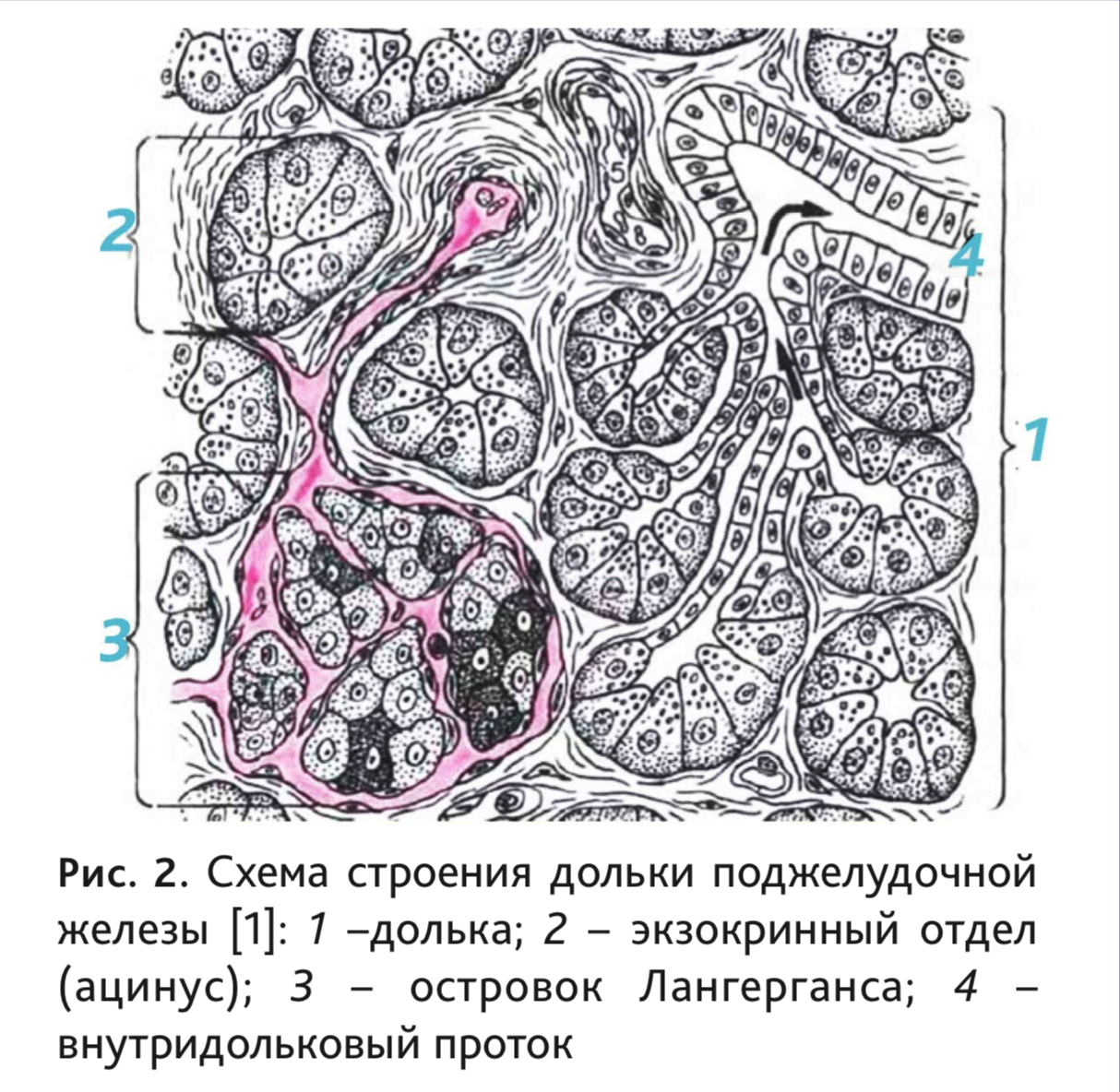
Экзокринная часть человека и рассматриваемых видов животных схожа по строению. Она
состоит из структурно-функциональных единиц – ацинусов, а также системы протоков, включающих
в себя вставочные, внутридольковые, междольковые протоки и общий панкреатический проток. Между
ацину-
сами лежат ретикулярные волокна, капилляры, нервные волокна и ганг-
лии.
Ацинус состоит из 8–12 экзокринных панкреоцитов или ациноцитов, лежащих на базальной мембране и нескольких протоковых (центроацинарных) эпителиоцитов, образующих вставочный проток.
Ациноциты выполняют секреторную функцию и вырабатывают пищеварительные ферменты. Они имеют пирамидальную форму. Широкое основание лежит на базальной мембране. Узкая апикальная часть имеет микроворсинки, выступающие в просветы ацинуса. По восприимчивости красителей клетку можно разделить на верхушечную часть, или зимогенную зону и базальную часть, или гомогенную зону. Зимогенная зона окрашивается кислыми красителями и является оксифильной. В ней можно различить крупные секреторные гранулы, в которых находятся ферменты в неактивном состоянии, при этом гранулы имеют различную плотность, что определяет их зрелость. В гомогенной зоне осуществляется синтез ферментов, поэтому она пронизана гранулярной эндоплазматической сетью с большим количеством рибосом, что обусловливает базофильное окрашивание. В нижней части клеток располагается ядро, также содержащее 1–2 ядрышка. Надъядерную область занимает комплекс Гольджи, рядом с которым находится большое количество митохондрий.
Вставочный проток образован уплощенными клетками со светлой цитоплазмой, окружающей овальное ядро. Они могут примыкать к ацинарным клеткам с боку или проникать в центр ацинуса. Вставочный проток продолжается межацинозными протоками (ductus interacinosi), а затем переходит во внутридольковые протоки (ductus intralobulares). Стенки протоков выстланы однослойным кубическим эпителием, а вокруг них расположена рыхлая соединительная ткань с кровеносными капиллярами и нервами.
Внутридольковые протоки собираются в междольковые протоки (ductus interlobulares), которые лежат в междольковых перегородках и впадают в общий проток поджелудочной железы, который открывается в двенадцатиперстную кишку. Междольковые протоки и общий проток покрыты слизистой оболочкой, представленной высоким призматическим эпителием с небольшим количеством бокаловидных клеток и эндокриноцитов, которые вырабатывают панкреозимин и холецистокинин. Они лежат на собственной пластинке слизистой оболочки, в которой можно обнаружить мелкие слизистые железы [2, 11, 30, 36, 41].
Эндокринная часть представлена панкреатическими островками (insulae pancreaticae), или островками Лангерганса, которые лежат между ацинусами (рис. 3). У крыс клетки эндокринной ткани составляют менее 2% [11], у человека их количество не превышает 3% [2]. У человека они имеют округлую или овальную форму, но могут быть и лентовидными и звездчатыми [2]. Число, размеры, форма и микроскопическое строение островков Лангерганса не одинаковы у разных позвоночных [5], а их диаметр колеблется от 50 до 500 мкм [42].
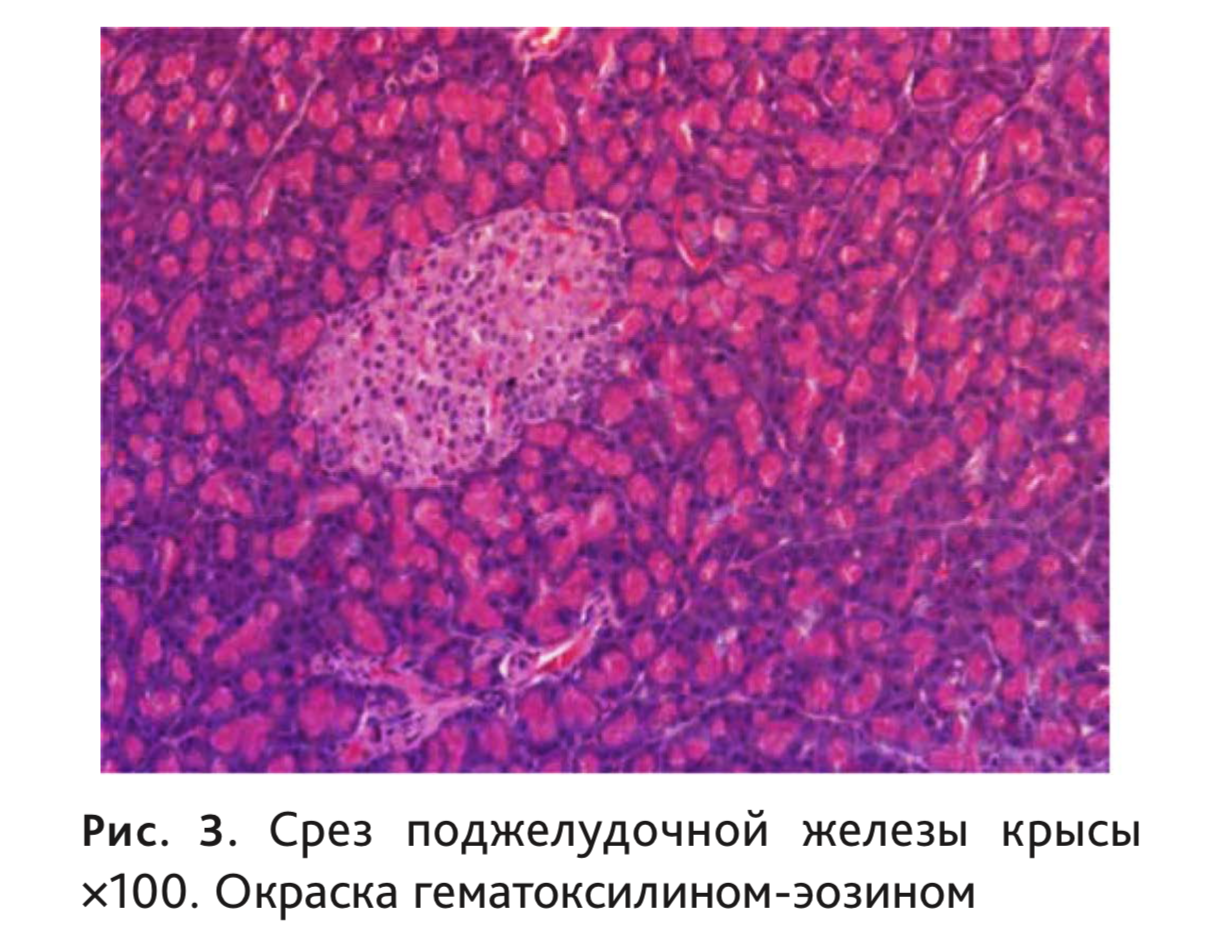
Состоят островки из инсулоцитов, вокруг которых лежат фенестрированные капилляры с перикапиллярным пространством, куда поступают синтезируемые гормоны. Капиллярная сеть островков сходна с клубочковой системой почки и имеет от 1 до 3 афферентных артериол. Плотность сосудов здесь в 5 раз выше, чем в экзокринной части.
Инсулоциты – небольшие клетки со светлой цитоплазмой, хорошо развитым аппаратом Гольджи, большим количеством митохондрий и секреторных гранул. Различают следующие виды клеток: базофильные (B-клетки), ацидофильные (A-клетки), дендритические (D-клетки) и РР-клетки. При светооптической микроскопии строение клеток у животных и человека схоже, но при электронно-микроскопическом исследовании можно обнаружить более выраженную видовую специфичность [11, 43] в распределении, размере и плотности гранул и органел. Так, например, у крыс секреторные гранулы В-клеток округлые с равномерной плотностью, а у морских свинок они могут быть полиморфными – дольчатыми, подковообразными различной плотности. У морских свинок средний диаметр А-гранул – 180 нм, В-гранул – 230 нм, а у кроликов размер А-гранул – 200 нм, В-гранул – 230 нм [43]. Кроме того, в панкреатических островках кроликов присутствуют фибриллярные структуры, не обнаруживающиеся в островковых клетках животных других видов [43]. Однако изучение ультраструктуры поджелудочной железы заслуживает отдельного рассмотрения. В данном обзоре подробно описать это невозможно.
A-клетки, на долю которых и у человека и у животных приходится до 25% всех клеток [2, 24, 44],
имеют обычно неправильную форму и окрашиваются кислыми красителями в красный цвет. Их плотно расположенные
в центре гранулы, отделенные от мембраны узким светлым ободком, содержат контринсулярный гормон –
глюкагон. Характерной особенностью А-клеток является наличие в их цитоплазме многочисленных секреторных гранул,
представляющих собой округлые тельца высокой электронной плотности, окруженные четко различимой гладкой мембраной.
Между плотным содержимым секреторной гранулы и ее отграничивающей мембраной имеется узкая электроннопрозрачная
зона (ореол). Электронноплотное «ядро» гранул обычно правильной округлой формы, его содержимое имеет
мелкозернистый вид. Ядро А-клеток – округлое, расположено несколько эксцентрично. Митохондрии имеют
удлиненную палочковидную форму и извилистые контуры. В цитоплазме некоторых А-клеток иногда имеется
небольшое количество гранул гликоге-
на [42].
Основную массу клеток (у человека и у животных, по разным источникам, от 75 до 80% [2, 24, 44]) составляют B-клетки, которые окрашиваются базофильными красителями, а их гранулы содержат инсулин. Они обычно полигональной или неправильной формы, и являются наиболее крупными по размерам. Эндоплазматический ретикулум клеток развит хорошо, он образован короткими канальцами, а также небольшими вакуолями, на мембранах которых находятся связанные рибосомы. Имеется также значительное число свободных рибосом, расположенных поодиночке или небольшими группами между клеточными органеллами. Все это придает цитоплазме В-клеток более «темный» вид по сравнению с А-клетками. Удлиненные митохондрии многочисленны и сосредоточены в основном вблизи комплекса Гольджи, который развит хорошо и занимает большую площадь клетки. В-инсулоциты могут быть представлены «светлыми» клетками, в цитоплазме которых мало гранул, имеющих разные размеры и плотность. Они обычно распределены по цитоплазме или сконцентрированы в одном из полюсов. Органеллы представлены единичными цистернами гранулярного эндоплазматического ретикулума, элементами комплекса Гольджи и мелкими удлиненными митохондриями. «Темные» В-клетки содержат большое количество гранул разных размеров и интенсивности окраски, которые равномерно расположены в цитоплазме. В этих клетках органелл мало, в основном обнаружены мелкие митохондрии и несколько компактно расположенных мелких липидных включений. 3-й тип клеток, определяемый, как «промежуточный», имеет равномерное распределение среднего количества гранул, типичный набор элементов органелл, мелкие липидные включения, вакуоли. Среди 3 типов В-клеток можно встретить клетки с явными признаками деструкции: резкая вакуолизация цитоплазмы, иногда вытесняющие цитоплазму липидные включения [42].
D-клетки, количеством до 10% [2, 24, 44], в панкреатических островках встречаются довольно редко, они расположены поодиночке или небольшими группами по всему островку, имеют полигональную или звездчатую форму и длинные цитоплазматические отростки. D-клетки или непосредственно контактируют с капиллярами, или посылают к ним свои отростки. Характерной особенностью D-клеток является наличие в цитоплазме специфических секреторных гранул, размер которых значительно варьирует. Наибольшее скопление гранул наблюдается в цитоплазматических отростках и на сосудистом полюсе. Ядро D-клеток – крупное, имеет округлую или несколько вытянутую форму и содержит компактное ядрышко. Содержимое гранул – соматостатин, который обладает способностью тормозить секрецию инсулина и глюкагона, a также ограничивать всасывание глюкозы в кишечнике. Это влияние соматостатин может осуществлять паракринно, действуя как местный гормон [42].
Менее 1% полигональных мелких РР-клеток вырабатывают панкреатический полипептид. Их можно обнаружить по периферии островков, лежащих в области головки железы; единичные клетки определяются также в экзокринной части железы [2, 24, 44]. Они содержат в своей цитоплазме очень мелкие секреторные гранулы (80 нм). Последние располагаются обычно в 1 ряд вдоль плазматической мембраны [42].
Кроме того, существуют промежуточные или ациноостровковые клетки, которые располагаются среди экзокринной ткани вблизи панкреатических островков и характеризуются особо высокой активностью окислительных энзимов цикла Кребса – сукцинатдегидрогеназы, НАДН-дегидрогеназы, высокими показателями энергетического и белкового обмена. Эти клетки имеют ультраструктурные признаки, свойственные эндокринному и экзокринному типам клеток, одновременно синтезируя гормоны и пищеварительные ферменты. Особенность этих клеток с промежуточной экзокринно-эндокринной морфологией – наличие гранул – крупных зимогенных и мелких, характерных для островковых клеток, которые имеют разный гормональный профиль. Они содержат эндоплазматический ретикулум 2 типов – шероховатый и гладкий, митохондрии – крупные, свойственные экзокринным клеткам, и мелкие, типичные для островковых клеток. Экзокринные гранулы этих клеток располагаются в апикальных отделах, и их содержимое выделяется в просвет центрацинозных протоков. Эндокринные гранулы находятся в примыкающих к капиллярам базальных отделах, и их содержимое попадает непосредственно в кровь. По представлениям целого ряда исследователей, при определенных функциональных состояниях ациноостровковые клетки могут переходить в ацинарные или островковые [45].
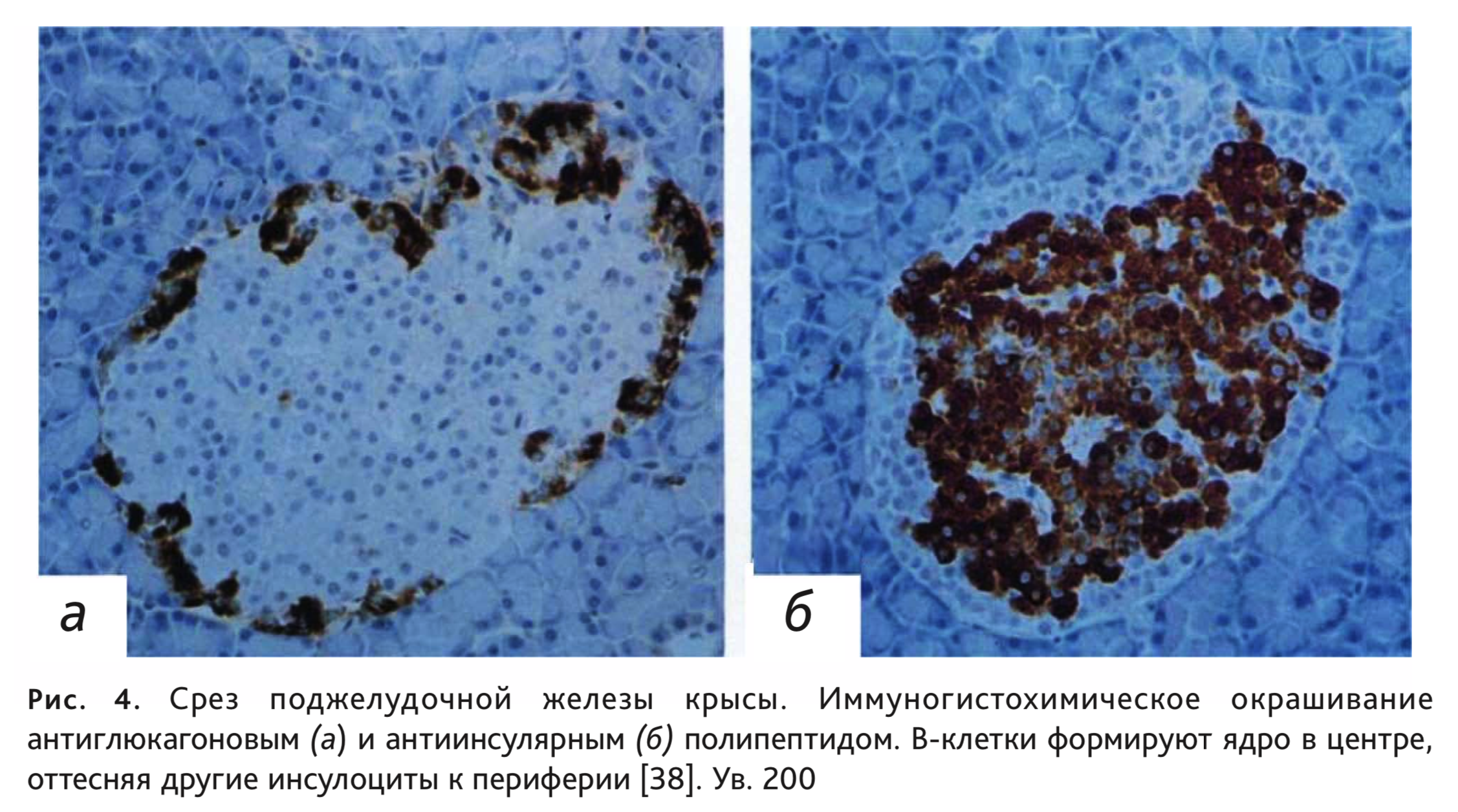
Клеточный состав островков в поджелудочной железе имеет свои видовые отличия. У человека количество разных типов инсулоцитов распределяется как по островкам, так и внутри островка преимущественно равномерно, а у грызунов наблюдается разделение островков по преобладающим А- или В-клеткам, но чаще всего В-клетки, располагаясь в центре, формируют ядро, вокруг которого А-клетки образуют мантию с включением небольших групп и единичных клеток других типов (рис. 4). У морских свинок центральные В-клетки оттесняют другие клетки к периферии, но в центре можно выявить наличие групп D-клеток. Островковый аппарат кроликов отличается высокой гетерогенностью: островки могут состоять полностью из В-клеток или А-клеток, возможно также преобладание 1 типа клеток (чаще – В, но бывает даже D-тип), причем расположение их тоже разнообразно и вероятно, как центральное положение популяции инсулоцитов, так и смещение к периферии или полюсу островка [24, 44].
Кровоток в поджелудочной железе имеет свои особенности, он направлен от островков к ацинарной ткани. Структура микроциркуляторного русла представлена островково-экзокринной портальной системой. Прекапиллярные артериолы направляются к панкреатическим островкам, которые первыми получают артериальную кровь. Сразу же после вступления артериола распадается на капилляры, которые образуют своеобразный сосудистый клубочек. Кровь, оттекающая из островков и содержащая высокую концентрацию гормонов, попадает в капиллярную сеть экзокринной ткани поджелудочной железы, а не непосредственно в панкреатические вены, что обеспечивает возможность влияния этих гормонов на экзокринную паренхиму. Капиллярная внутриостровковая сеть имеет постоянное (векторное) направление кровотока от инсулин- к глюкагон- и далее – к соматостатин-секретирующим клеткам. Наличие инсулоацинарной портальной системы является одной из основ функционального взаимодействия экзокринной и эндокринной функций поджелудочной железы [23, 45, 46].
У человека капилляры имеют собственную мембрану, которая контактирует с мембраной инсулоцитов, формируя таким образом, перикапиллярное пространство. У мышей есть только одна – общая мембрана [36], но нет разделяющей базальной мембраны между панкреатическими островками и панкреатическими ацинусами. Между эндокринными и экзокринными клетками наблюдаются соединения в виде десмосом, интердигитаций, плотных и щелевидных контактов. На этом основании можно предположить, что клетки связаны друг с другом структурно и функционально [45].
Установлено, что регенеративные процессы тканей поджелудочной железы происходят не равномерно по всей ткани, а определёнными участками и в разное время, причем обновление в пределах каждой дольки происходит одновременно и в эндокринном, и в экзокринном отделах [9].
Самообновление в поджелудочной железе возможно при участии перибилиарных желез, располагающихся в стенке крупных желчных протоков или около них и открывающихся в их просвет. Установлено, что, помимо продукции муцина, в перибилиарных железах обнаружены ниши мультипотентных стволовых клеток, которые могут дифференцироваться в гепатоциты, холангиоциты и клетки островков поджелудочной железы. Перибилиарные железы внепеченочных желчных протоков описаны у человека, лабораторных животных (крыс, мышей), а также у большинства млекопитающих, но они не обнаружены они пока только у рептилий [47].
Заключение
Строение поджелудочной железы человека и лабораторных животных имеет много общего, как в анатомическом, так и гистологическом строении.
Источник развития и закономерности органо- и гистогенеза поджелудочной железы человека и животных схожи, но различаются сроками. Образовываясь из энто- и мезодермы, она имеет несколько зачатков: 2 – у человека и 3 – у грызунов. Однако вопрос органогенеза пока недостаточно изучен и характеризуется противоречивыми данными ряда авторов.
Наиболее яркое отличие имеет анатомическое строение железы, поскольку в зависимости от формы выделяют несколько типов строения органа: компактный, рассеянный (брыжеечный) и промежуточный. У человека поджелудочная железа компактная. Большинство авторов относит поджелудочную железу рассматриваемых животных к брыжеечному типу, но существуют мнения, что у хомяков строение органа компактное, у крыс и морских свинок смешанного типа, а у кроликов имеет место вариативность – возможен как брыжеечный тип, так и компактный. Несмотря на разнообразие формы у каждого вида в железе выделяют головку, тело и хвост.
Протоки поджелудочной железы также крайне разнообразны. У человека хорошо изучены варианты анатомического строения и преимущественно наблюдают 2 протока – главный и добавочный, которые открываются сосочками в двенадцатиперстную кишку. У животных, видимо, имеет место большая вариативность, поэтому описание протоков у разных авторов отличается. Так, у крыс выделяют протоки, которые впадают в желчный проток или сразу в двенадцатиперстную кишку, по другим данным, имеется всего 1 проток. У мышей описаны несколько дополнительных протоков открывающихся мелкими сосочками. Особенностью кроликов является значительное разобщение желчных протоков и протоков поджелудочной железы, но данные о количестве добавочных протоков весьма противоречивы.
Гистологическое строение железы также однотипно, значительную специфичность можно обнаружить только на ультрамикроскопическом уровне. Однако в данном обзоре это подробно не рассматривалось.
Экзокринная часть не имеет существенных различий. А в строении островкового аппарата можно выделить видовые различия в качественном и количественном составе инсулоцитов в островках Лангерганса. Так, например, у грызунов, в отличие от человека, присутствует разделение островков по преобладающим А- или В-клеткам, но чаще всего В-клетки, располагаясь в центре, формируя ядро, окруженное А-клетками и небольшими группами или единичными клетками других типов.
Список источников
- Ильиных М.А. Структурно-функциональная характеристика поджелудочной железы потомства самок крыс с хроническим поражением гепатобилиарной системы различного генеза. Автореф. дисс. на соиск. уч. степ. канд. биол. наук, Челябинск. 2007: 20.
- Афанасьев Ю.И., Юрина Н.А. Гистология, цитология и эмбриология. М.:Медицина.2002: 744.
- Бектимирова М.Р. Морфофункциональное состояние поджелудочной железы и уровень общего белка в сыворотке крови крыс при введении полиоксидония. Scientific notes. 2017, 232:12–7.
- Сулаева О.Н. Учебное пособие по гистологии. Запорожский государственный медицинский университет. 2015:13.
- Яковлева О.В., Ситдикова Г.Ф., Яковлев А.В. Практикум по эндокринологии. Методическое пособие. Казань, 2007: 26.
- Luis Rodrigo, Janeza Trdine Rijeka. Acute Pancreatitis. Croatia InTech. 2011: 288.
- Петренко В.М. Поджелудочная железа в эмбриогенезе. Бюлл. науки и практики. 2017, 6: 72–89.
- Пивченко П.Г., Пивченко Т.П. Сравнительный анализ развития поджелудочной железы у человека и белой крысы в эмбриональном периоде (обоснование экспериментальной модели). Оригинальные научные публикации. 2016: 80–3.
- Калигин М.С., Титова А.А., Плюшкина А.С., Титова М.А., Гумерова А.А., Киясов А.П. Пролиферация клеток поджелудочной железы при экспериментальном диабете. Гены & клетки. 2014, Том 4, 3: 85–8.
- Денисов С.Д., Пивченко Т.П. Динамика структурных изменений поджелудочной железы в эмбриогенезе белой крысы. Современные аспекты фундаментальной и прикладной морфологии: сборник трудов научно-практической конференции, посвященной 110-летию со дня рождения академика НАН Беларуси Д. М. Голуба. Минск: БГМУ. 2011: 316.
- Minoru Tsuchitani, Junko Sato Hiroko Kokoshima A comparison of the anatomical structure of the pancreas in experimental animals. Toxicol Pathol. 2016. Vol 29: 147–54.
- Сапин М. Р. Анатомия человека в двух томах. М.:Медицина. 2001.Т. 2: 640.
- Петренко В. М. Лимфоузлы поджелудочной железы у морской свинки. Международный журнал прикладных и фундаментальных исследований. 2014. 4: 52–4.
- Петренко В.М. Форма и топография поджелудочной железы у крысы. Успехи современного естествознания. 2012. 2: 35–9.
- Петренко В.М. Форма и топография поджелудочной железы у морской свинки. Международный журнал прикладных и фундаментальных исследований. 2013. 7: 36–9.
- Andreas Hornblad. Imaging the Pancreas New Aspects on Lobular Development and Adult Constitution. Print & Media Umeo. 2011: 50.
- Thomas Nolte, Patricia Brander-Weber, Charles Dangler. Nonproliferative and Proliferative Lesions of the Gastrointestinal Tract, Pancreas and Salivary Glands of the Rat and Mouse. Toxicol Pathol. 2016: 62
- Макарова М.Н., Рыбакова А.В., Гущин Я.А., Шедько В.В, Мужикян А.А., Макаров В.Г. Анатомо-физиологическая характеристика пищеварительного тракта у человека и лабораторных животных. Международный вестник ветеринарии. 2016. 1: 82–104.
- Петренко В.М. Форма и топография поджелудочной железы. Международный журнал прикладных и фундаментальных исследований. 2015. 1: 1114–7.
- Lesa Thompson. Pet rabbit nutrition – structure and function of its gastrointestinal tract. Veterinary Times. 2013. 44(17): 12–4.
- Ron Rees Davies, Jennifer A.E. Rabbit gastrointestinal physiology. Vet Clin Exot Anim. 2003. 6: 139–53.
- Rosen Stefanov Dimitrov. Comparative Ultrasonographic, Anatomotopographic and Macromorphometric Study of the Spleen and Pancreas in Rabbit (Oryctolagus cuniculus). Not Sci Biol. 2012. 4(3):14–20.
- Fraser P. A. Henderson J. R. The Arrangement of Endocrine and Exocrine Pancreatic Microcirculation Observed in the Living Rabbit. Quarterly Journal of Experiment Physiology. 1980. 65:151–8.
- Steiner D.J., Kim A., Miller K., Hara M. M.Pancreatic islet plasticity: interspecies comparison of islet architecture and composition. Islets. 2010. 2(3): 135.
- Жеденов В. Н. Анатомия кролика. М.: Сов. наука. 1957: 312.
- Yunan Wang, Abudurexiti Kayoumu, Guotao Lu, Pengfei Xu, Xu Qiu, Liye Chen, Rong Qi, Shouxiong Huang, Weiqin Li, Yuhui. Experimental Models in Syrian Golden Hamster Replicate Human Acute Pancreatitis. Wang & George Liu. 2016: 9.
- Cheryl L. A practical guide to the histology of the mouse. Scudamore Mary Lyon Centre, MRC Harwell. 2013: 248.
- Привес М. Г., Лысенков Н. К., Бушкович В. И. Анатомия человека: учебник. – 12-е изд., испр. и доп. СПб. : Гиппократ. 2008: 704.
- Kapil Vashisht, Sherry L. Nady, Rita D. Engler, Brain K. Kelsch, Susan N. Lynk, Benjamin R. Cape, Guenter Hoffmann, Emily K. Meseck, Robert C. Jonson. Extraparenchymal Bile. Pancreatic Ducts and Duodenal Papillae: Pathologic Evaluation in Nonclinical Species - A Brief Review. Toxicologic Pathology. 2015. 43: 651–61.
- Suckow M. A., Karla A. S., Ronald P. W. The laboratory rabbit, guinea pig, hamster, and other rodents. American College of Laboratory Animal Medicine series. Amsterdam; Boston: Elsevier Academic Press. 2012: 1261
- Юдичев Ю.Ф., Дегтярев В.В., Гончаров А.Г.Анатомия животных: учебное пособие. В 2-х т. Т. 2 . Оренбург: Издательский центр ОГАУ. 2013: 406.
- Cathy A. Johnson-Delaney. Anatomy and Physiology of the Rabbit and Rodent Gastrointestinal System. Proceedings of Association of Avian Veterinarians, Association of Exotic Mammal Veterinarians (AEMV) Sessions. 2006: 9–17.
- Ноздрачев А. Д., Поляков Е. Л. Анатомия крысы. СПб.: Лань. 2001: 464.
- Reznik G., Reznik-Schüller H., Mohr U. Clinical Anatomy of the European Hamster: Cricetus cricetus. L.U.S. Government Printing Office, Washington. 1978: 251
- Wagner J.E., Manning P. J. The Biology of the Guinea Pig. American College of Laboratory. Animal Medicine Academic Press. 1976: 317
- Treuting P. M., Dintzis S.M., Frevert C.W. Comparative Anatomy and Histology: A Mouse and Human Atlas. Elsevier, Amster-dam-Boston-Heilderberg-London-NewYork-Oxford-Paris-SanDiego-SanFrancisco-Singapore-Sydney-Tokyo. 2012: 461
- Proshchina A.E., Krivova Y.S., Barabanov V.M., Saveliev S.V. Ontogeny of neuro-insular complexes and islets innervation in the human pancreas. Front Endocrinol (Lausanne). 2014. 5: 1–8.
- Szczurkowski A., Kuchinka J., Nowak E., Kuder T. Autonomic innervation of pancreas in Egyptian spiny mouse (Acomys cahirinus, Desmarest). Acta Veterinaria Brno. 2009. 78: 557–61.
- Andrada Seicean. Challenges in Pancreatic Pathology. InTechOpen. 2017: 240.
- Федорова Н.Н., Сентюрова Л. Г. Влияние этанола на клетки поджелудочной железы белых мышей. Вестник АГТУ. 2010. 1(49): 59-60.
- Eurell J. A., Frappier B. L. Dellmann's Textbook of Veterinary Histology. Sixth edition.Blackwell Publishing. 2006: 416.
- Целуйко С.С., Красавина Н.П., Корнеева Л.С., Лашин С.А., Остронков В.С. Морфофункциональная характеристика поджелудочной железы и легкого при экспериментальной гипергликемии на фоне применения дигидрокверцитина. Благовещенск. 2017: 151.
- Кирилловых А.С. Морфометрическая характеристика эндокринной части поджелудочной железы грызунов при остром деструктивном панкреатите. «Инновации студентов в области ветеринарной медицины». 2011: 38–43.
- Islam M. S. The Islets of Langerhans. Springer Science & Business Media. 2010: 798.
- Баранов С.А., Нечаев В.М. Поджелудочная железа как единый функционально взаимосвязанный орган. Эндокринология. Медицинский совет. 2017. 11: 148–51.
- Arun R. Pandiri, Eric Schultze. Overview of the Pancreatic Toxicity and Carcinogenesis Session. Pancreatic Toxicity and Carcinogenesis. Toxicologic Pathology. 2014. 42: 204–6.
- Шапирова Э.И., Газизов И.М., Гумерова А.А., Киясов А.П. Перибилиарные железы желчных протоков как ниша мультипотентных стволовых клеток. Гены & клетки. 2014.Том IX, 3: 29–33.
Поиск



